引言
提起中生代,很多人脑中立刻就会想起那些高大威猛的恐龙。作为一类家喻户晓的古生物,恐龙在中生代长达一亿多年的霸业总是为人津津乐道,而它们在白垩纪末大灭绝中的遗憾退场也总是令人惋惜。好在,还是有一支恐龙的后裔——鸟类得以幸存,并在新生代重新繁盛了起来,直至今日。
但是,有一类生物与恐龙的命运非常相似,却鲜有人知。不知道你是否有过这么一个问题:白垩纪($\text{Cretaceous}$)作为中生代最后一个纪元,它到底为什么叫这个名字呢?
其实无他,白垩($\text{chalk}$)是一种颜色雪白、内部多孔而极易粉碎的特殊石灰岩,这种岩石绝大多数都形成于这一纪元。在欧洲部分地区,这种岩石甚至可以在海边堆成高达上百米的悬崖,比如英吉利海峡边著名的多佛白崖,最高处约 $110$ 米,东西长度达到 $13$ 千米:

虽然看起来非常壮观,但白垩实际上是由海洋中浮游生物的遗骸构成的,只不过在漫长的地质变迁中,它们的肉体早已灰飞烟灭,唯有碳酸钙外壳被留下,进而密密麻麻地堆在一起形成了白垩。你或许很难想象,到底要有多少遗骸才能沉积成如此震撼人心的白色坟场;而这片坟场的主人,也正是这篇文章的主角——球石藻($\text{Coccolithophores}$,又名颗石藻)。
严格来说,白垩通常还包含其他海洋生物的钙质外壳,比如有孔虫(有孔虫门,$\text{Foraminifera}$),但球石藻占据了大部分。你或许从未听说过这个名字,但它们却是中生代最王道的藻类家族。它们中的大部分都会制造碳酸钙鳞片,并用这些鳞片做一身盔甲,把自己包在里面。据估计,在白垩纪末的最鼎盛时期,球石藻一度占到全体浮游藻类生物量的 $50\%$ 左右,可以说是碾压一切同行的存在:

(上图:$C.~pelagicus$,属球石藻科球石藻属($Coccolithus$),可能是最经典的球石藻形象)
(上图:翼球藻属($Alisphaera$)的 $A.~unicornis$)

(上图:三毛金藻属($Prymnesium$)的 $P.~parvum$,属于球石藻纲基部的普林藻目($\text{Prymnesiales}$),没有碳酸钙外壳)
球石藻纲($\text{Coccolithophyceae}$)又名普林藻纲($\text{Prymnesiophyceae}$),属于定鞭藻门($\text{Haptophyta}$)。如今,它们依然广泛分布于世界各地的水体之中(主要是海洋),但却已不复往日的繁荣。$6600$ 万年前的白垩纪末大灭绝对于整个定鞭藻门都是灭顶之灾,门内超过 $90\%$ 的物种直接绝灭。在新生代,球石藻虽然也有过短暂的回光返照,但最强藻类的宝座终究还是拱手让给了硅藻(硅藻纲,$\text{Bacillariophyceae}$),直至今日。
所以,在它们的背后,到底是一段怎样的演化历程呢?作为曾经的王者,它就真的这样自甘陨落了吗?
正文
序章:起源与早期演化
长期以来,人们都认为与定鞭藻亲缘关系最近的类群是中阳虫(中阳虫门,$\text{Centroheliozoa}$),二者共同组成定鞭界($\text{Haptista}$)这一大类群;但最近,许多研究都把另一个名为网鞭虫(网鞭虫门,$\text{Telonemia}$)的类群拉进了定鞭界,并认为它才是定鞭藻的姐妹。如此,色藻类的两大分支就变成了定鞭界与 $\ce{SAR}$ 类群:
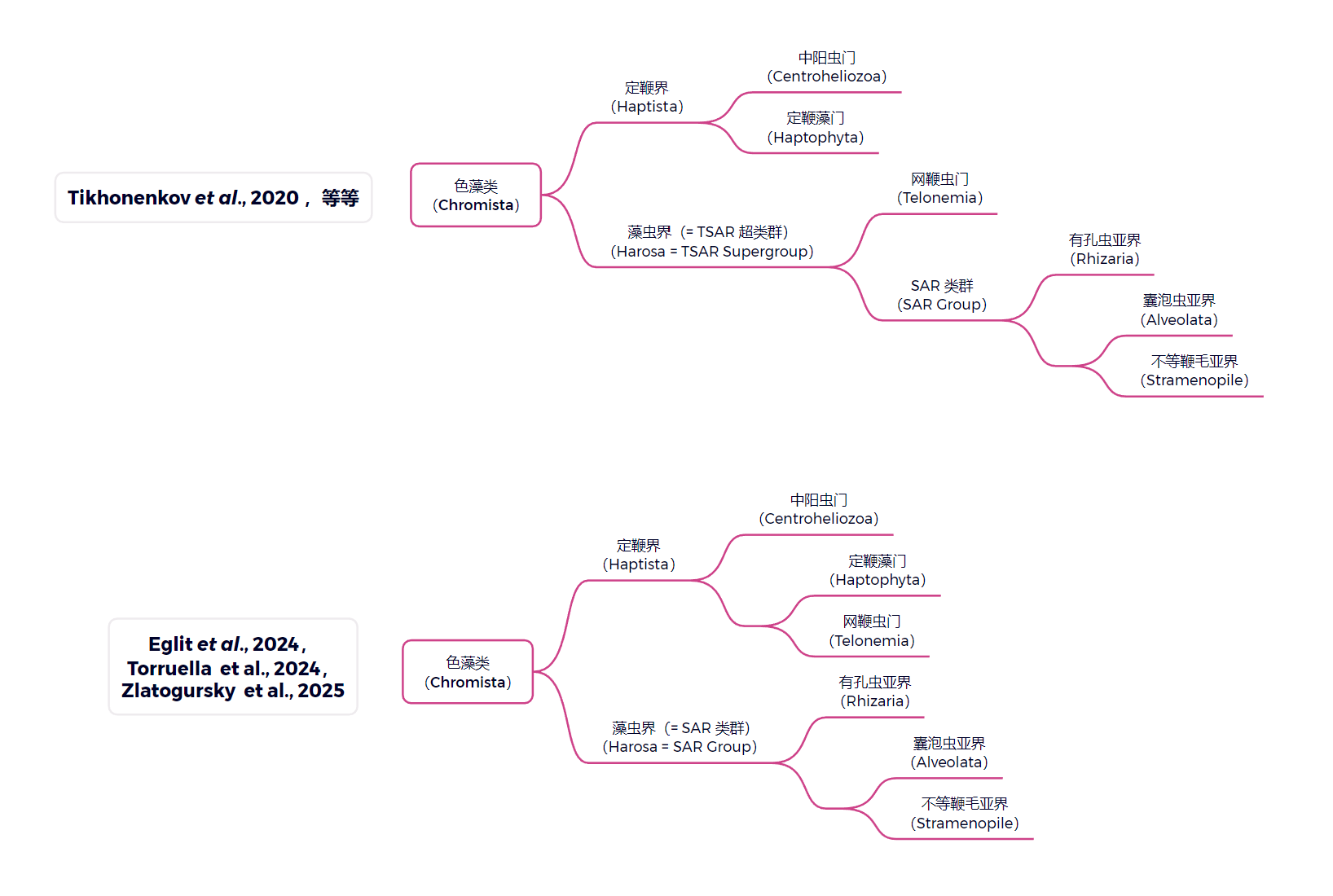
(上图:色藻类的演化树,请注意传统观点和新观点中网鞭虫的位置变化。一篇关于中阳虫的文章:Link)
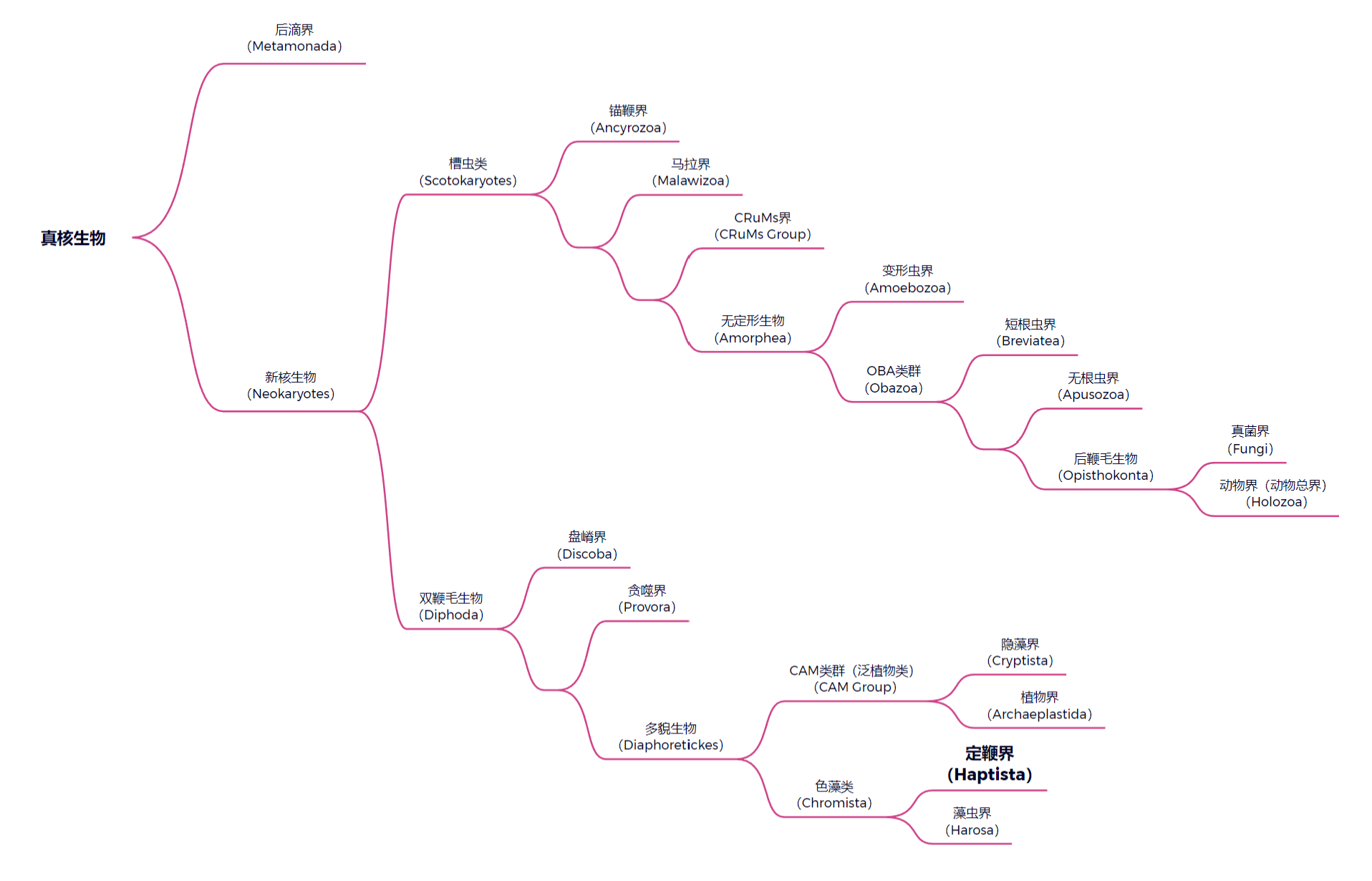
(上图:真核生物演化树中定鞭界的位置,基于 Torruella et al., 2024。需要注意,真核生物的系统发育树仍未完全确定,一些类群的位置和地位有争议。)
这样一来,网鞭虫很可能就代表了定鞭藻祖先的模样:拥有两根长袖善舞的鞭毛,自由运动,以吞噬细菌和比自己小的真核生物为生,是单细胞生物中不折不扣的凶猛掠食者:
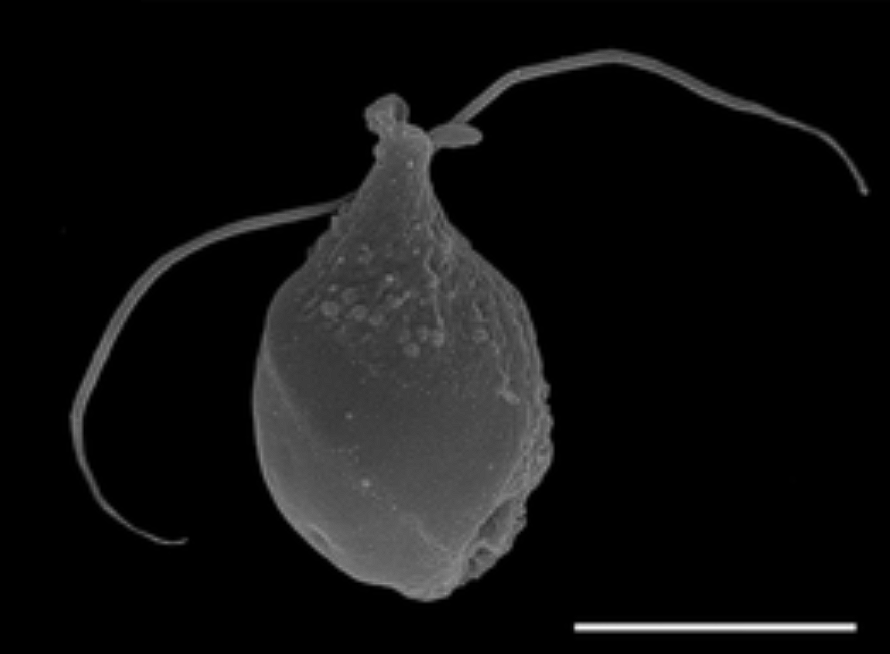
(上图:网鞭虫 $T.~rivulare$,长得像个运动水壶)
我们的故事要从 $5$ 亿多年前的寒武纪开始讲起。当此之时,四海之内遍布着各种真核藻类,而隐藻(隐藻门,$\text{Cryptophyta}$)正是其中较为繁盛的一支。它们的祖先在很久之前吞噬了一颗红藻(红藻门,$\text{Rhodophyta}$),而在某种机缘巧合之下使其内共生在了自己的细胞当中,从而成为了第二批真核生物光合作用者。如今,它们已经崛起成了海洋中制造氧气的一方诸侯,当然也顺理成章地成为了我们主角日常的食物之一。
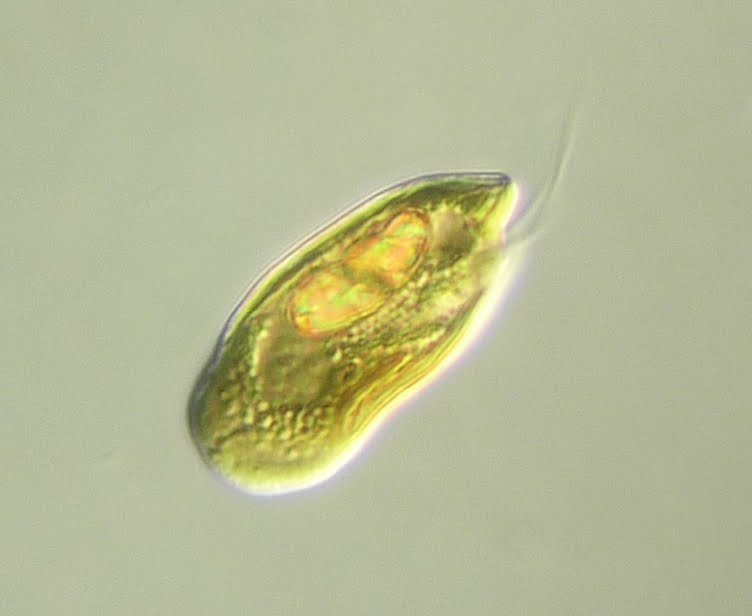
(上图:一只隐藻,具体物种不详)
接下来发生的事情,估计各位也可以猜到了。某一天,当定鞭藻的远古鼻祖吞下一颗隐藻之后,在某种机缘巧合之下并没有将其完全消化,尤其是那颗内共生的红藻被保留了下来。就这样,定鞭藻的祖先夺走了隐藻的质体,也获得了光合作用的能力,从此便正式转职为了藻类。
在刚开始,任凭谁都看不出它能有什么作为。毕竟,定鞭藻来的实在是太晚了,藻类的生态位早已被几大家族瓜分殆尽。休说绿藻、红藻,就连同为色藻类的亲戚——淡色藻(淡色藻门,$\text{Ochrophyta}$),都遥遥领先它几亿年。况且,即使是单论能力,它也没有什么过人之处,便只能在各路强权的夹缝中苟且偷生。

(上图:淡色藻门,如今最为繁盛的藻类门类,包括褐藻(左下)、硅藻(上中)、金藻(右)等)
唯一能拿得出手的,或许只有那个被称为“定鞭”($\text{haptonema}$)的特殊结构了。这玩意夹在两根鞭毛之间,在表面上与真的鞭毛没什么两样,但实际上结构(主要是微管的排列方式)非常不同。这使得定鞭非常灵活,什么盘绕、伸缩、卷曲都不在话下。在定鞭藻还是掠食者时,定鞭是它有力的猎杀工具,但现在似乎也没什么大用了。只能依靠它附着在基质上,游走于狭缝中,祈求一线生机。
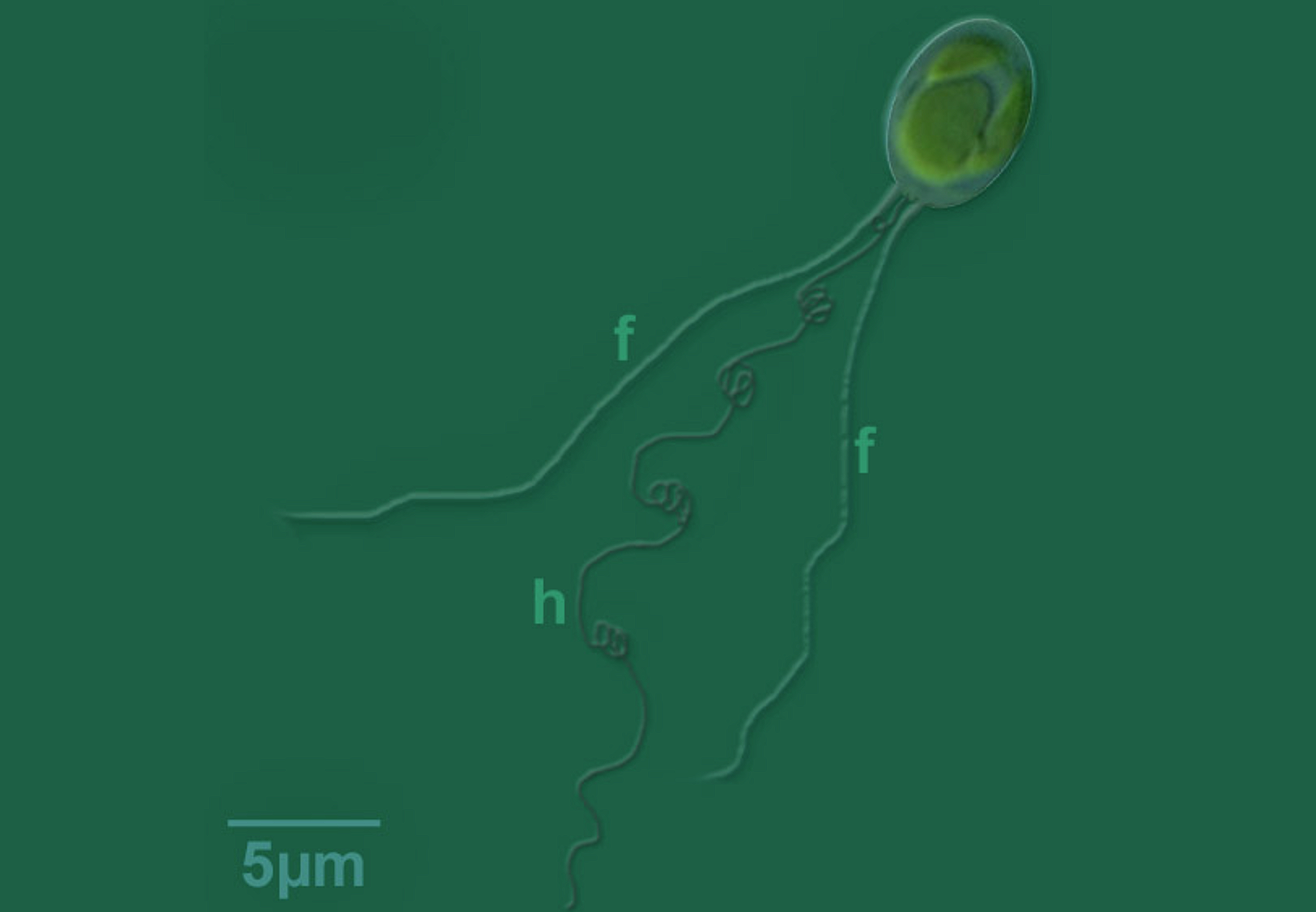
(上图:金色藻属的 $C.~polylepis$,拥有整个门中最长的定鞭(图中 $\text{h}$ 标注的结构),可以达到几十微米)
但是,无常的命运却突然给了它一份馈赠。在一次基因转移事件中,它意外获得了一份来自淡色藻的基因,而且不是别的,正是关于光合作用的基因。我们至今仍不知道这起事件的缘由是什么,但总之,定鞭藻从此学会了合成岩藻黄素($\text{fucoxanthin}$)。这种光合色素虽然只起辅助作用,但却是非常超模的存在,既能当“天线”收集阳光,又能在质体中高效地进行能量转移,同碳双烯的特殊结构还赋予了它强大的抗氧化活性。可以想象,在有了岩藻黄素后,定鞭藻的光合效率一定迈上了好几个台阶,并终于能和当时主流的藻类平起平坐。
也正是因此,许多定鞭藻在显微镜下都泛着金光,导致早期的生物学家错误地把它归为了金藻。这一错误直到 $21$ 世纪,分子生物学家出手才得以纠正。
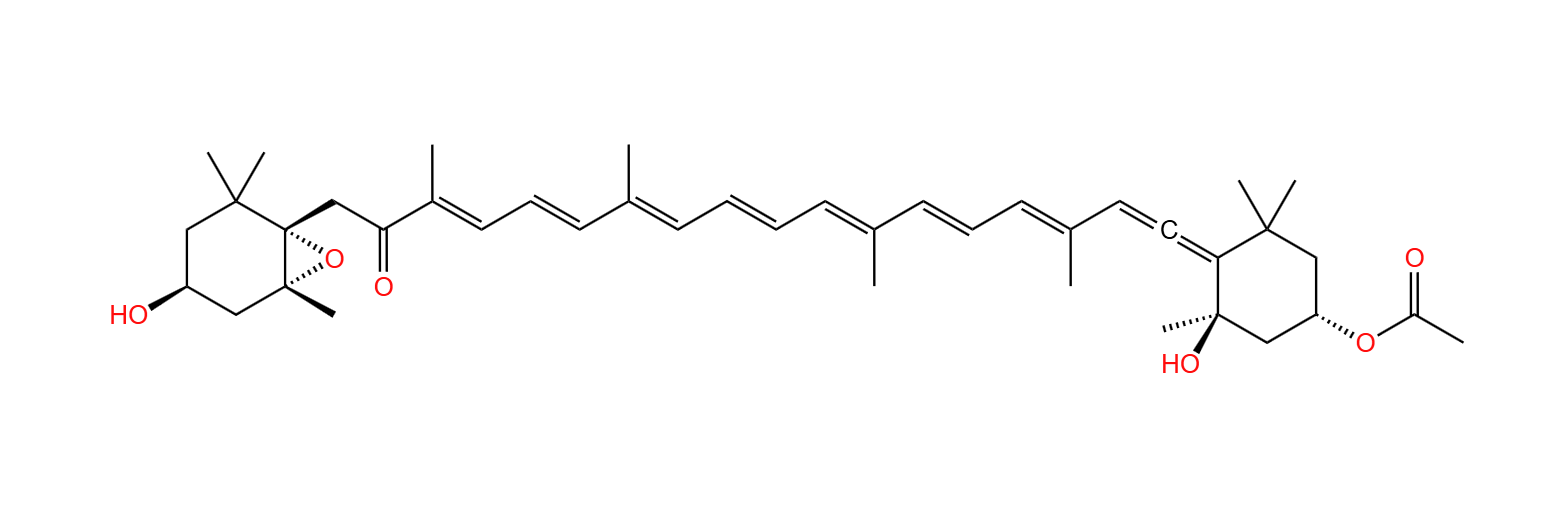
(上图:岩藻黄素的化学结构)
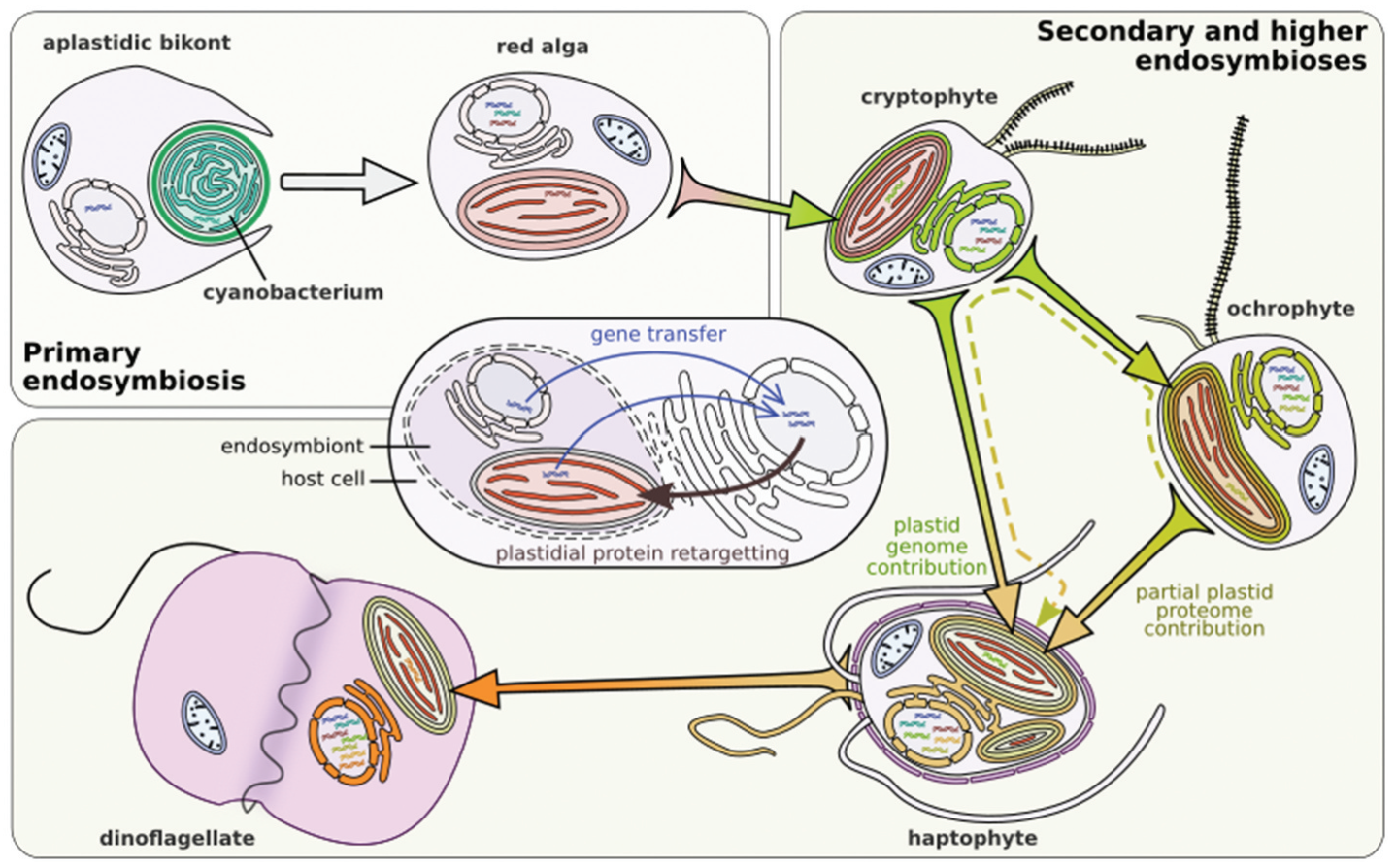
(上图:关于定鞭藻的一系列内共生和基因转移事件的总结)
就这样,定鞭藻终于——还是没有成功崛起。毕竟基因转移这种事就像抄作业,还是无法撼动先发者的优势。不过,这至少给了定鞭藻一些信心,或许它所要做的就是继续等下去,等到时间给它一个机会。
于是,在大约 $3.7-3.5$ 亿年前,地球海洋突然遭遇了一系列原因不明的灾厄,一会儿全球变冷海平面下降,一会儿又大缺氧乃至有毒物质暴增,海洋生物死伤无计,史称泥盆纪末大灭绝($\text{end-Devonian mass extinction}$)。但是,这样的灭绝事件对于定鞭藻来说却是极大的利好。可能是因为部分竞争对手的倒下,也可能是因为大量动植物的死亡造成的富营养化刺激了藻类的繁殖,总之在纪元之交,定鞭藻终于迎来了第一次多样化。如今门下三纲的祖先应该就是在那时登上了历史舞台:
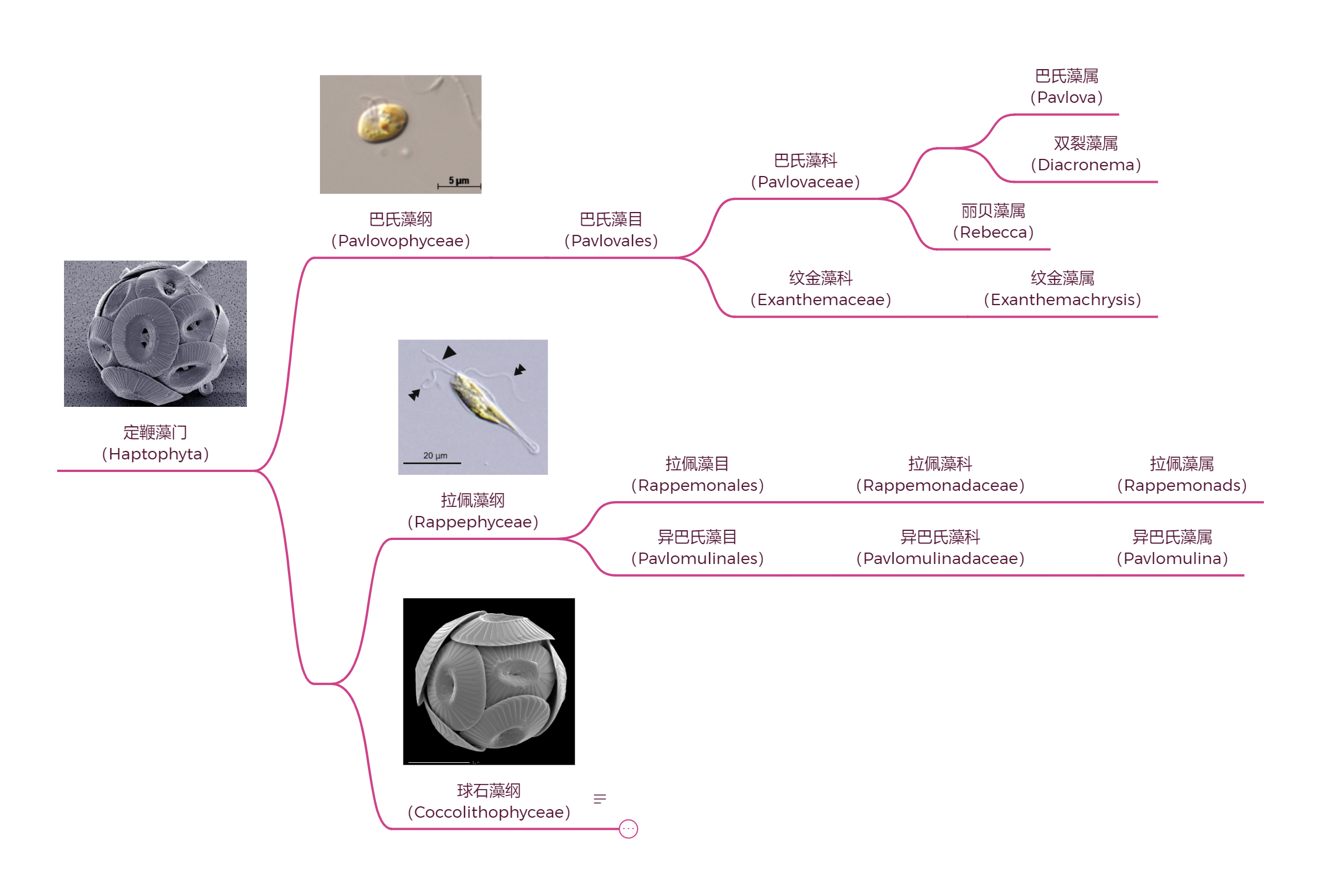
(上图:定鞭藻门基部谱系的系统发育。需要注意,关于巴氏藻和拉佩藻谁先分化是有争议的,此处采用 Kawachi et al., 2021 的观点;实际上,分化时间应当很接近;省略了很多未被正式描述的类群)
巴氏藻纲($\text{Pavlovophyceae}$)被认为是最早分化的谱系。它们与其他定鞭藻相比,有两点明显的不同:一是具有不等长的鞭毛,二是演化出了一个被称为眼点的结构,可以用于感知光线强弱。
位于纲基部的类群是纹金藻属($Exanthemachrysis$),有时也被当作单独的一科。它们是浅海的底栖藻类,利用自己的定鞭附着在基质上,而这样的生活方式很可能是从定鞭藻的共同祖先那里开始,流传了几亿年的光阴。当然,纹金藻也并非不敢创新。它们发展出了群落的生存策略,通过分泌粘液将分散的细胞团结起来,便能利用体型优势劝退单细胞掠食者;群落的整体行为尚不清楚,但群落的直径可以从几十微米到将近一毫米,以至于肉眼依稀可见:
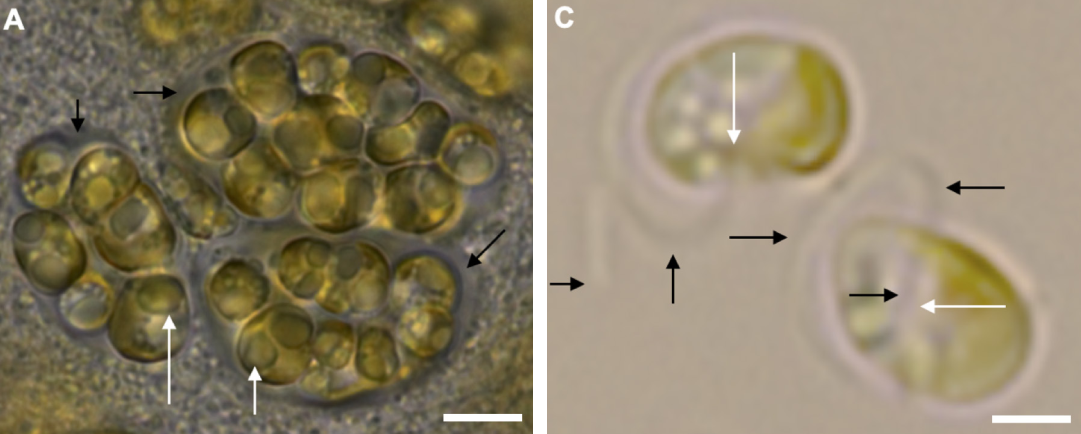
(上图:$E.~fresneliae$ 的群落(左)与单个细胞(右))
巴氏藻属($Pavlova$)则是一个全球性分布的大家族,其下有十几个物种。它的名字来源于俄罗斯著名芭蕾舞演员巴甫洛娃($\text{A.~Павловна}$),据说是因为这种藻类的运动方式像是跳芭蕾舞(这真的对吗?)。它有一根很长的前向鞭毛,在运动时会弯曲成 $\text{S}$ 形,从而带动整个细胞扭曲着前进:
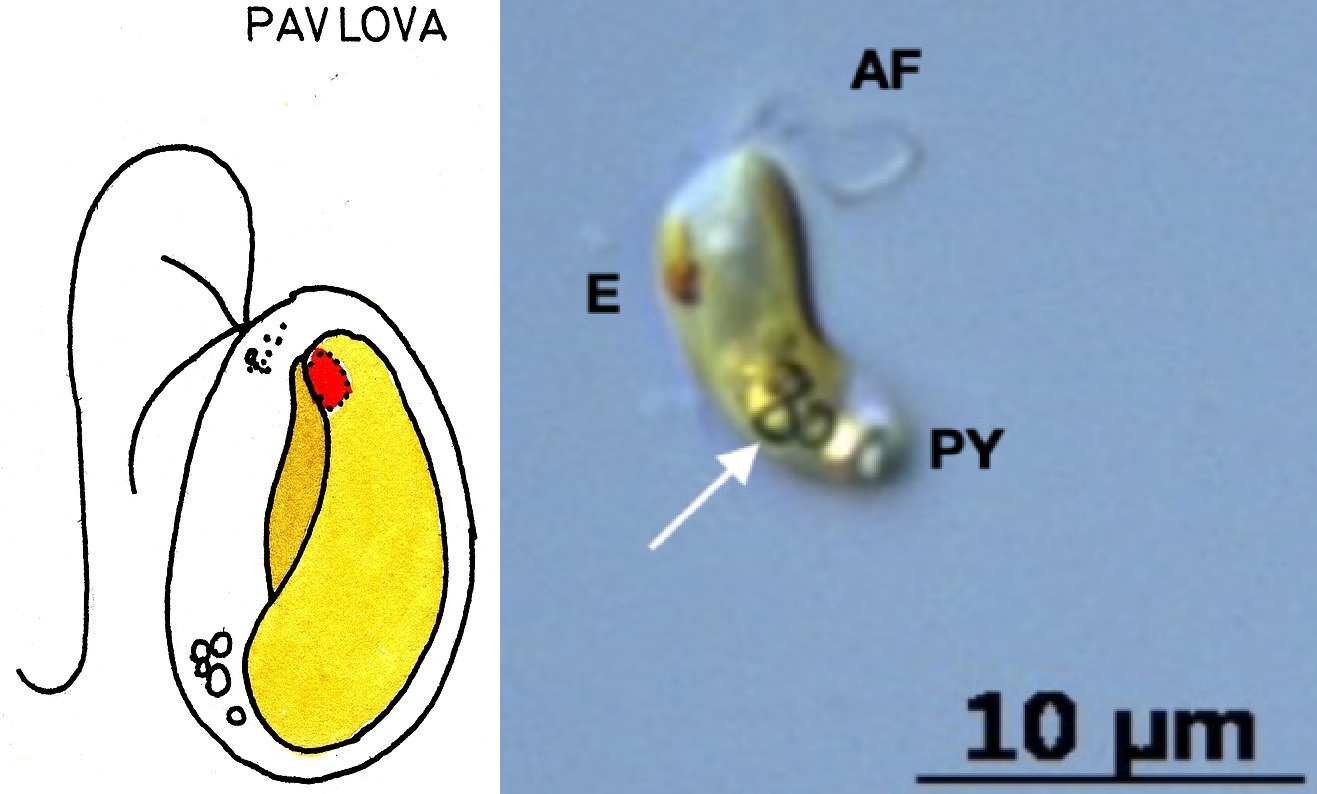
(上图:$Pavlova$ 属的简笔画与实拍(具体物种不明))

(上动图:$Pavlova$ 属的运动,画面很模糊,但依稀可见弯曲扭动的前鞭毛和较为明显的眼点;具体物种不详)
拉佩藻纲($\text{Rappephyceae}$)则是 $2021$ 年才根据分子生物学证据成立的类群,其中的拉佩藻目($\text{Rappemonales}$)完全是根据宏基因组建立,所以也没什么可讲。异巴氏藻($Pavlomulina$)就比较有意思了,它的外形像条小鱼,尺寸明显比其他定鞭藻大,体内有整整四个质体(其他定鞭藻一般只有 $1-2$ 个);身体几乎两侧对称,细胞后方还有一个明显的“尾巴”;两条鞭毛等长,定鞭突出指向前方。这些配置都表明了它对快速定向运动的适应:
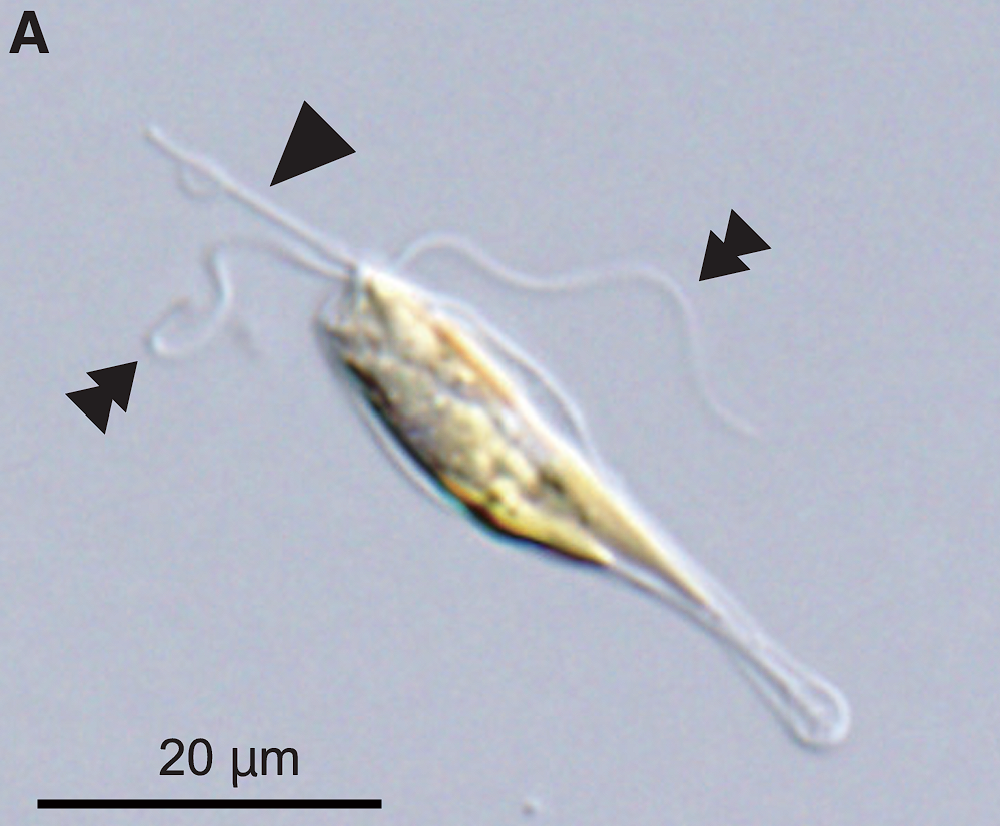
(上图:$P.~ranunculiformis$,图中双箭头指向鞭毛,单箭头指向定鞭)
如果你觉得上面这些花里胡哨的都没啥用,那至少有一支定鞭藻开始认真打磨起了自己的核心能力,那便是在序言里就被提到的球石藻纲($\text{Coccolithophyceae}$)。上文中,一段来自淡色藻类的基因让定鞭藻学会了合成岩藻黄素,而球石藻就选择在此基础上继续改进。最终,它们获得了以 $19’-$岩藻黄素醇己酸酯为代表的一系列衍生物。简单来说,这就像是以岩藻黄素为模板进行数值调整,比如耐久 $+10\%$、效率 $-10\%$ 等等。由于结构差别不大,这些衍生物之间的转化也很容易。于是,球石藻便能灵活控制胞内各种光合色素的浓度,大大提升自己对不同环境的的适应能力。
不过,球石藻显然认为这还不够。就在几天前,一篇登上 $\ce{Science}$ 期刊封面的文章破解了它们的另一项绝技:一种被称为岩藻黄素-叶绿素超级复合物($\text{fucoxanthin chlorophyll supercomplex}$)的光能天线。简单来说,它们会将岩藻黄素、叶绿素等多种光合色素负载到一些特定的蛋白质上,从而组装出一台巨大的分子机器。这台机器捕获光能的效率高达 $95\%$,约为陆地植物的三到四倍,因此也极有可能是已知最大、最复杂、效率最高的光合复合物:
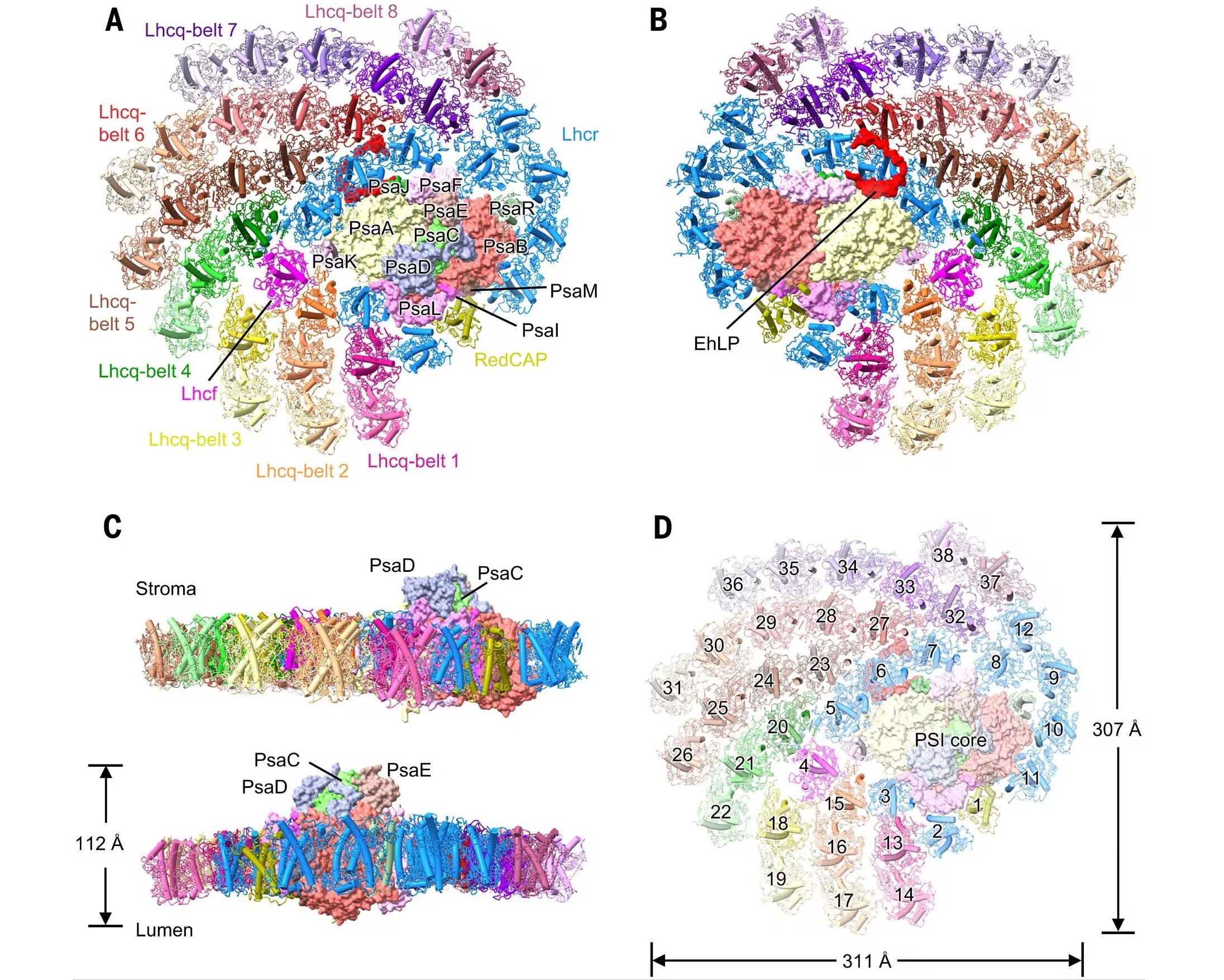
(上图:球石藻光合复合物的三维结构及其尺寸)
作为主角光环的持有者,冥冥中似乎一切皆有天意。当球石藻完成这一系列革新之后,地球进入了二叠纪($\text{Permian}$)。而很快,在距今 $2.5$ 亿年前,随着西伯利亚超级火山的一声巨响,地球迎来了显生宙以来最为惨烈的二叠纪末大灭绝($\text{end-Permian mass extinction}$)。以红藻、绿藻为代表的第一代真核藻类遭受重创,从此便跌下神坛,丧失了最主要浮游藻类的地位;与之相随的,便是次级内共生藻类开始全面崛起。在这股潮流中,球石藻倚仗着新的技能快速壮大,终于成为了定鞭藻中独秀的一枝,也让它有了冲击藻类王者宝座的野心。
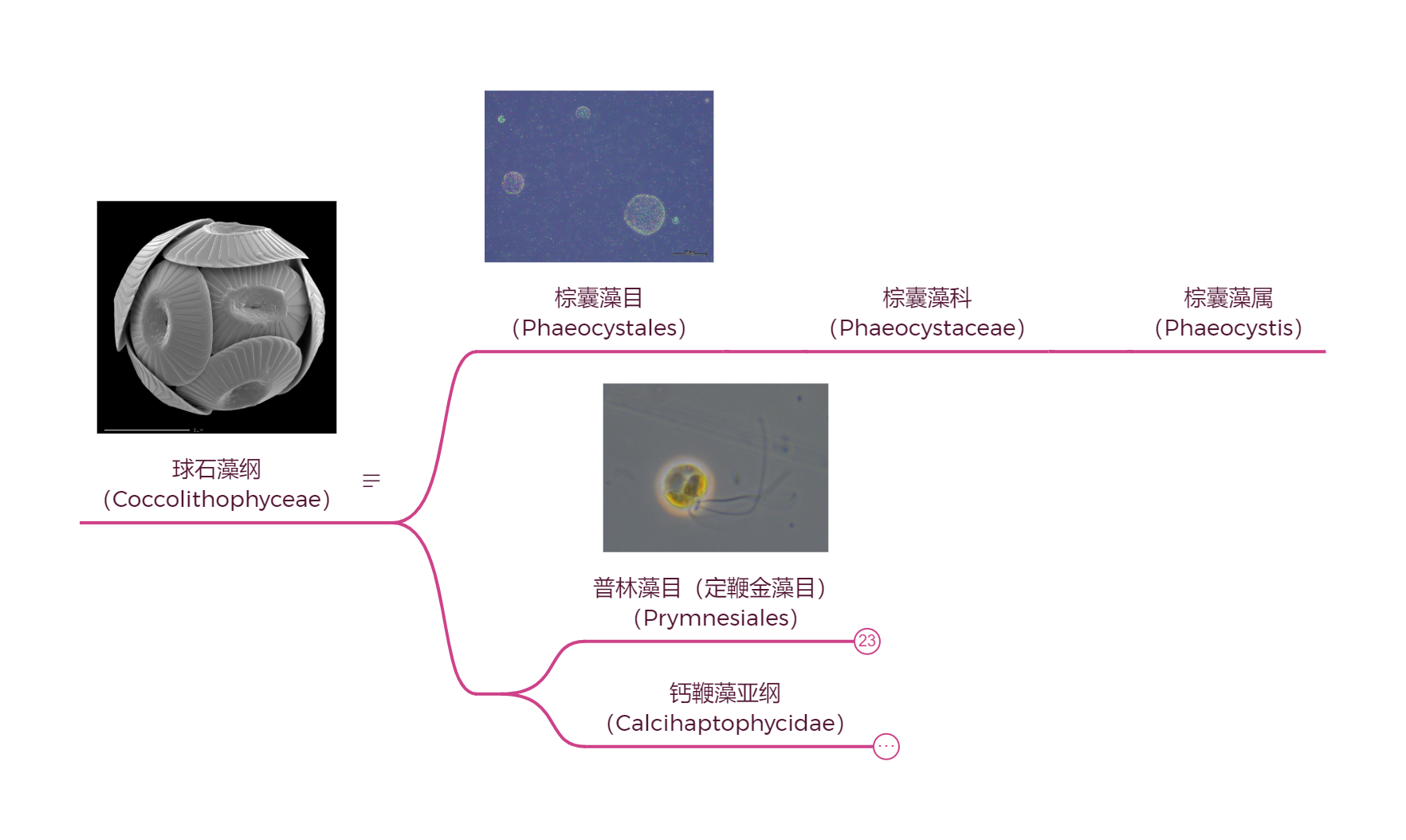
(上图:球石藻纲基部谱系的系统发育)
早期的球石藻并没有我们印象中标配的钙制铠甲,这点从位于纲基部的棕囊藻($Phaeocystis$)身上就能看出来(事实上,棕囊藻目的分化时间可能在二叠纪末大灭绝之前)。它的单个细胞平平无奇(至少从外表上看),但可以上百个细胞聚在一起形成群落。群落呈规整的球形,所有成员都被嵌在由多糖凝胶构成的透明基质中;直径一般在百微米级,但也能经常能找到毫米级甚至接近一厘米的大群落:

(上图:黑背景下的棕囊藻群落)
这些群落将终生在海洋中漂浮,随波逐流,宛如一座座晶莹的城堡。如果你运气非常好,还有机会见到一堆聚在一起的群落,即群落群;当然,还有罕见的独立棕囊藻细胞:
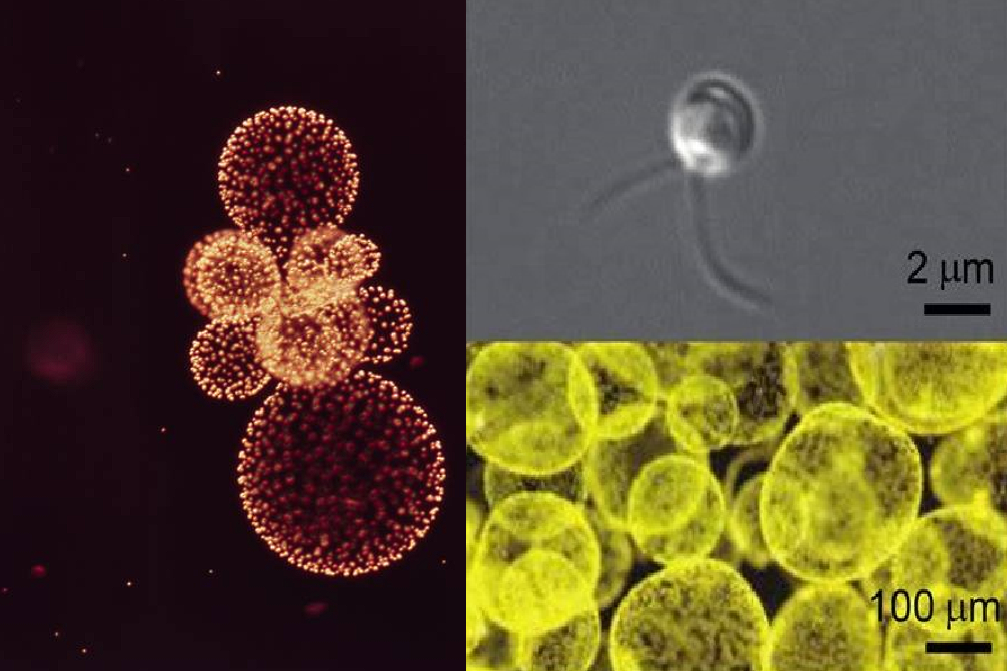
(上图左:一个棕囊藻群落群;右上:$P.~antarctica$ 的独立细胞;右下:培养皿中的大量棕囊藻群落)
但是,如果你运气不好,见到的棕囊藻可能就是下面这样的:海边飞扬的泡沫。在某些地区,每年的固定时节都会形成棕囊藻的藻华(主要是春夏季),由于群落中大量的多糖胶质,当海面被风浪搅动时,就会形成大量的气泡,从而形成这种奇特的景象:

(上图:摄于法国贝桑港,$2023$ 年 $6$ 月 $8$ 日;藻华形成物种可能是 $P.~globosa$)
一般来说,你并不会觉得这很浪漫,因为泡沫中的有机成分会发酵产生刺鼻的臭味,让人感到恶心;藻华也会扰乱食物链,从而影响当地的整个生态系统。正是因此,很多人都将棕囊藻归类为有害藻类。
普林藻目($\text{Prymnesiales}$),也被译为定鞭金藻目,是球石藻纲基部另一个没有钙质铠甲的类群。但是,这并不代表它们就不叠甲了,许多物种的体表还是覆盖着有机质鳞片。目下至少有 $13$ 个属,系统发育树如下(很多属找不到确切位置):
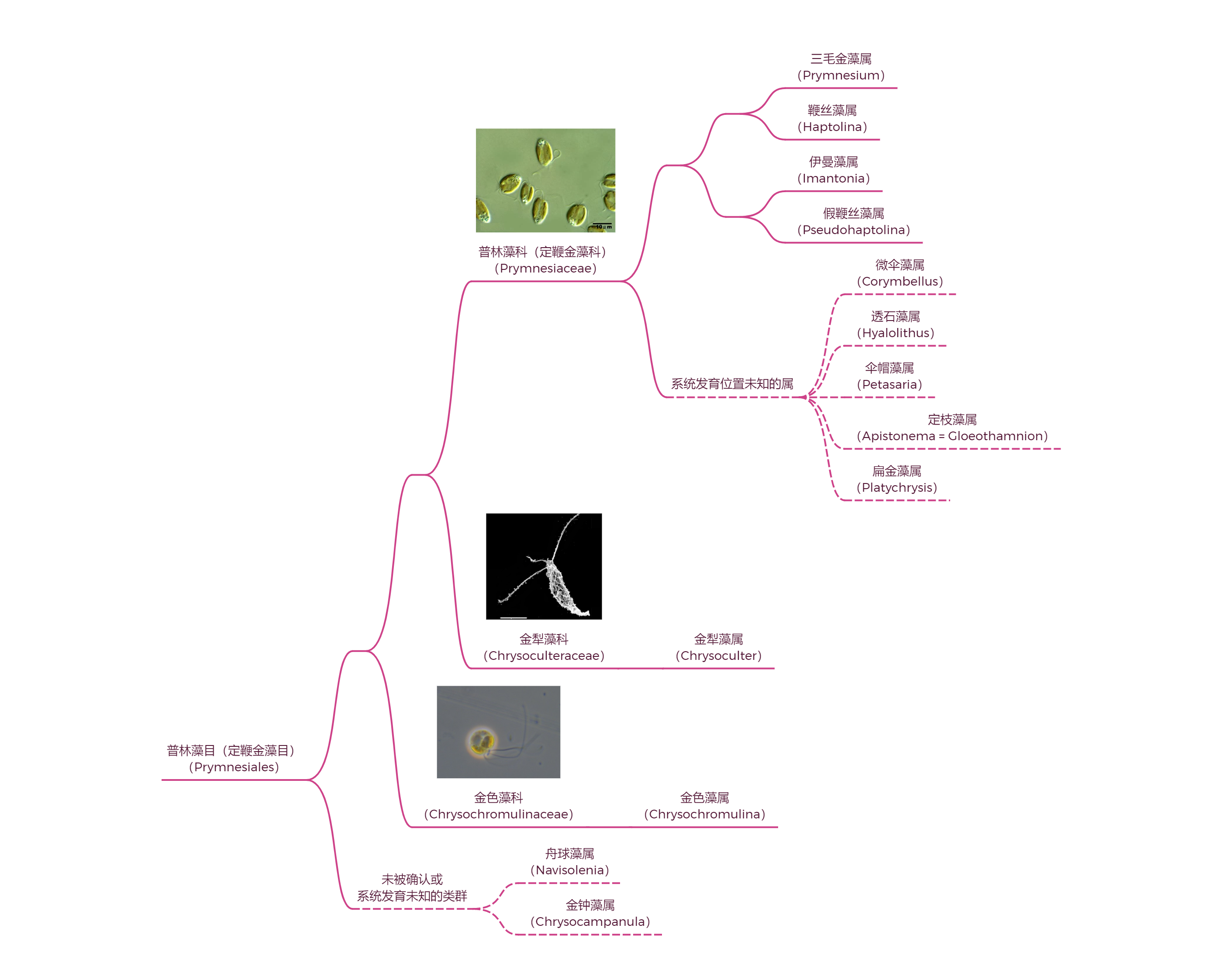
位于基干位置的金色藻($Chrysochromulina$)广泛分布于全球海域,虽然形态上很普通(也被认为比较原始),但却是个不折不扣的狠角色。每当它形成藻华之时,往往会造成大量其他生物死亡,甚至导致长时间的食物网崩溃。这是因为它演化出了某种致命的毒素(具体结构未知,但分子式似乎是 $\ce{C67H127ClO27}$),可以有效地杀死鱼类等大型海洋生物。比如 $1988$ 年春夏之交,在挪威附近海域就出现了严重的金色藻藻华,导致了近千吨养殖鱼类的死亡;此次事件持续影响着当地的生态系统,直到 $5$ 年后才得以完全恢复。
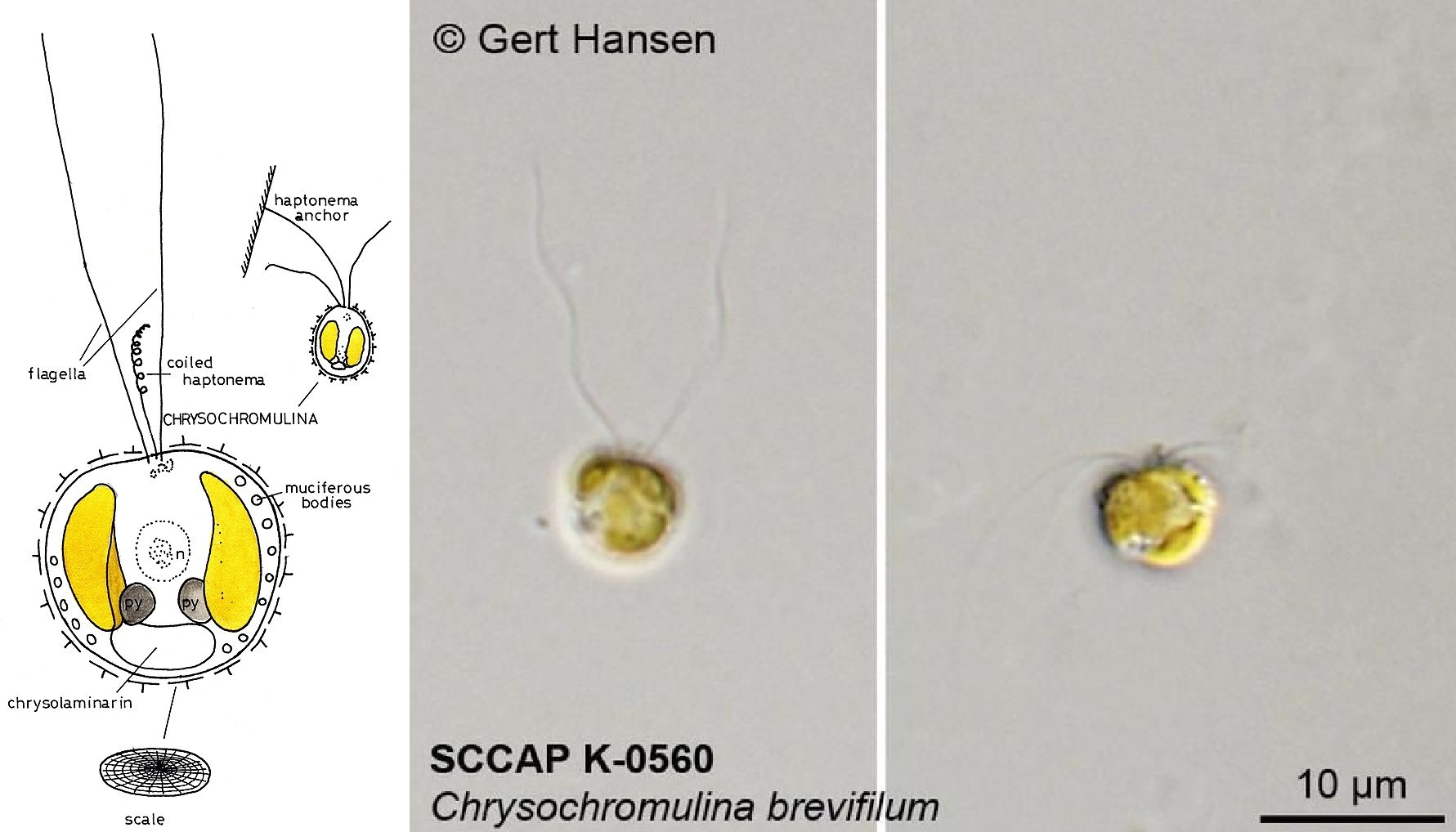
(上图:金色藻的简笔画与 $C.~brevifilum$ 实拍)
同样易形成有毒藻华的是三毛金藻($Prymnesium$)。这个名字其实是从很久之前流传下来的,实际上并不准确,但也就习惯成自然了。它是广盐性的藻类,在淡水和海洋中都能生长,对于温度和 $\ce{pH}$ 值也没有太高要求,因此十分常见;尤其在我国,还是生物防治的重点对象之一。
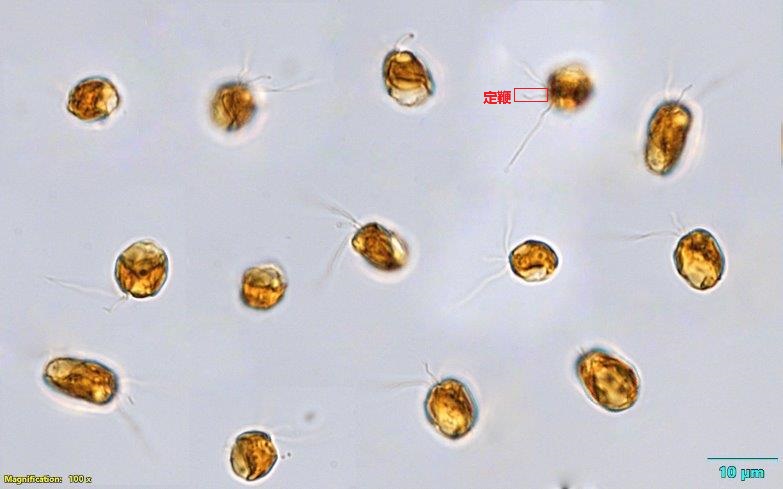
(上图:$P.~parvum$。用红框框出了一个个体的定鞭)
三毛金藻的毒素已经被人类破译,其中最主要的一种名为 $\text{Prymnesin-1}$,化学式为 $\ce{C107H154Cl3NO44}$,结构如下图所示。值得注意的是在去年,人们发现负责制造这种毒素的是一种名为 $\text{PKZILLA-1}$ 的巨型蛋白质,由 $45212$ 个氨基酸残基构成,也是目前已知最大的天然蛋白质。
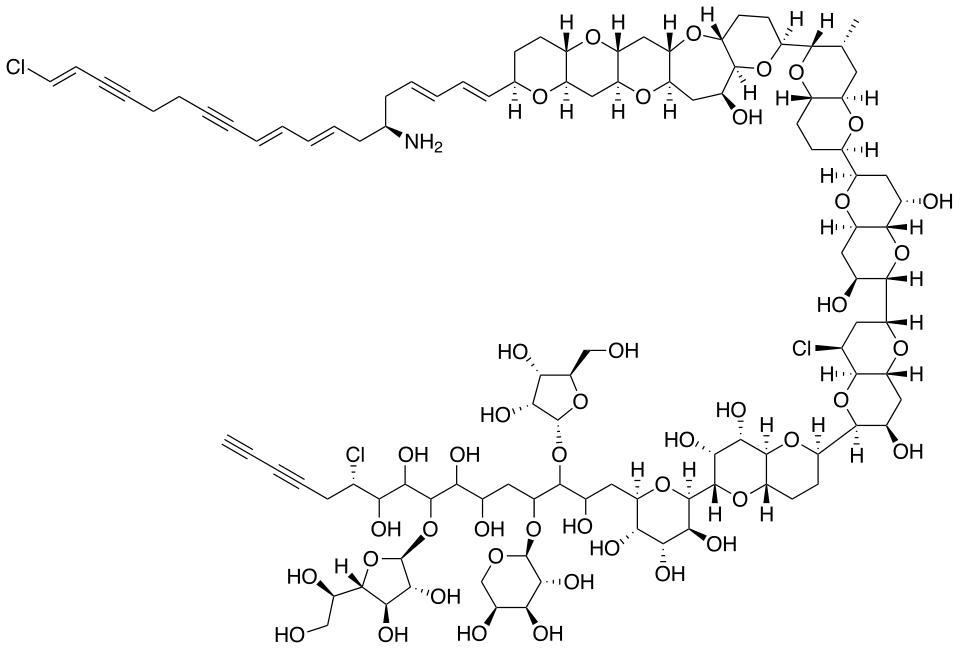
(上图:$\text{Prymnesin-1}$ 的化学结构)
金犁藻($Chrysoculter$)则可能是这一家子中叠甲叠得最厚的。它用两层有机质鳞片将细胞完全覆盖,外层的鳞片呈菱形,互相之间紧密贴合;内层鳞片呈小椭圆形,主要用来堵住外层的缝隙。整体外形像一把弯刀,或者用来犁地的锄头,绝对称得上是一朵奇葩:
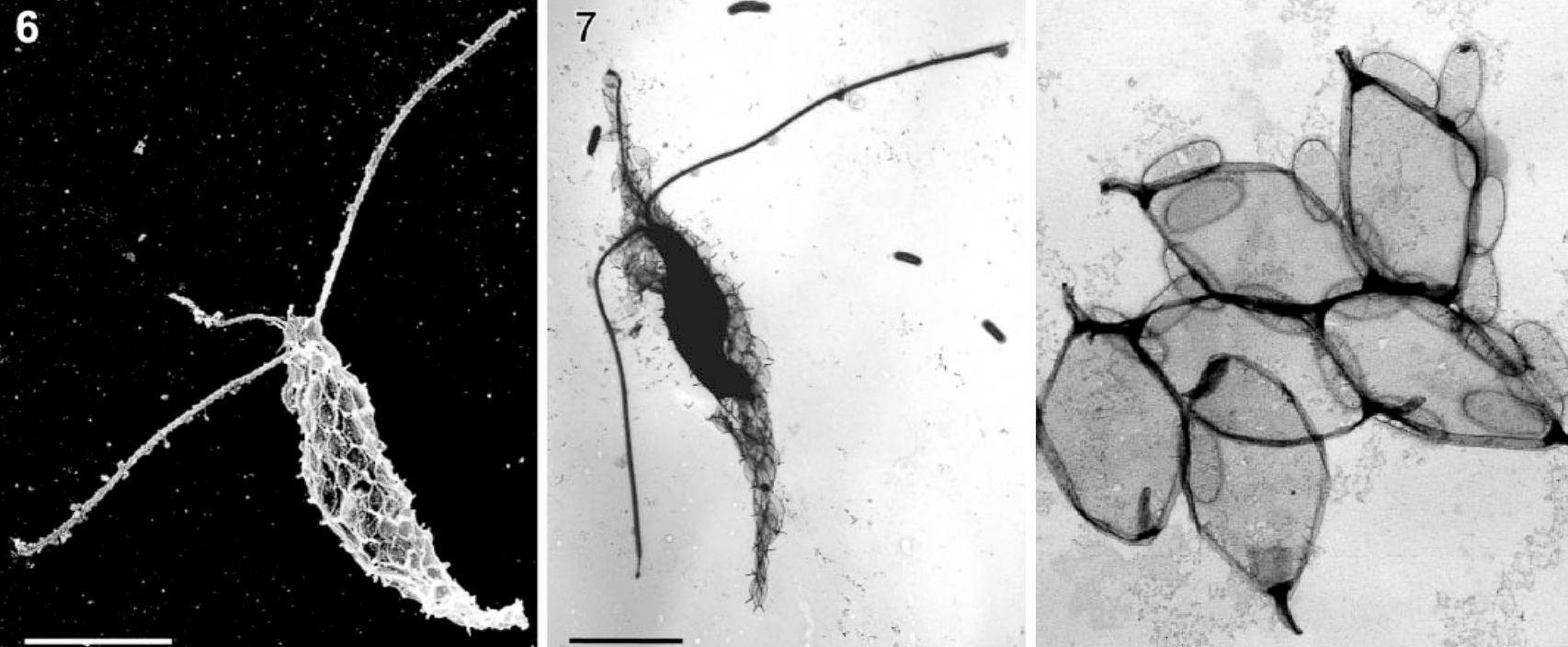
(上图:$C.~rhomboideus$ 与其鳞片特写)
不过,在这一大家子中,最奇妙的可能还得属定枝藻($Apistonema$)。它的身体形态与其他所有定鞭藻都迥然不同,简单来说,它已不再是浮游藻类。定枝藻将成百上千个细胞连为一体,形成了树枝状分叉的外观,整个群体也被底部细胞的定鞭牢牢地固定在基质上。这些特征都表明,定枝藻已经成功演化为了多细胞藻类(虽然体型还是比较小)。当然,在这条道路上,目前的它显然还无法与褐藻(褐藻纲,$\text{Phaeophyceae}$)竞争,所以也只能躲在沼泽湿地之类的地方猥琐发育,但若待到千万年之后,它又是否会作为后浪,扛起向褐藻类反攻的大纛呢?
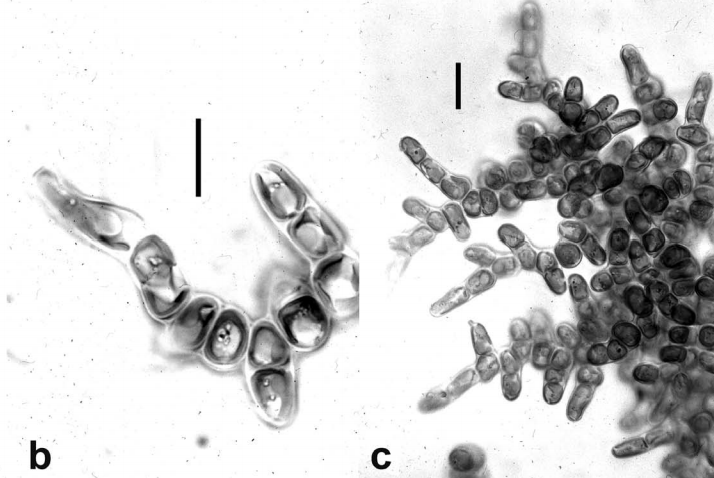
(上图:$A.~pyrenigerum$)
好吧,以普遍理性而言,这更可能会是一次无疾而终的演化尝试,毕竟普林藻目本就不是球石藻纲的主力。自三叠纪以来,家族里挑大梁的便一直是那个身披钙质铠甲的类群——钙鞭藻亚纲($\text{Calcihaptophycidae}$)。它们才是符合我们印象的、狭义的球石藻。
中章:盛世与毁灭
事实上,我们至今仍说不清楚它们为什么会钙化,因为有一点是毫无疑问的:为了制造足以覆盖全身的碳酸钙鳞片(专业点说叫“球石粒”,$\text{coccoliths}$),钙鞭藻设计了一套极为复杂的系统,而运转这套系统需要耗费大量的能量。
首先,它需要将足量的底物($\ce{Ca^{2+}}$ 与 $\ce{HCO3-}$)转运至细胞内,一个被称为球石囊泡的特殊结构中。在那里,碳酸钙会以 $\ce{Ca^2+ + 2HCO3- ->[酶] CaCO3 + CO2 +H2O}$ 的反应生成。根据计算,光这一步就大概要花费它们光合作用所获得的能量的三成。随后,碳酸钙晶体会以一些特殊的蛋白质为模板开始生长。在这一阶段,细胞内的高尔基体还会合成一些特殊的多糖(球石调节多糖,$\text{CAPs}$),它们就宛如一把把美工刀,在晶体的表面精雕细琢,以一种人类还无法理解的方式将碳酸钙“雕刻”为所需的形状。如此造就的鳞片无论是从精细程度还是从美观性上讲都是一件巧夺天工的艺术品,每次细细观之都不禁令人感叹。最后,球石囊泡会将完工的鳞片运送至细胞表面,多块鳞片之间彼此交叠,终成完全覆盖细胞的铠甲,即所谓“球石层”($\text{coccosphere}$)。
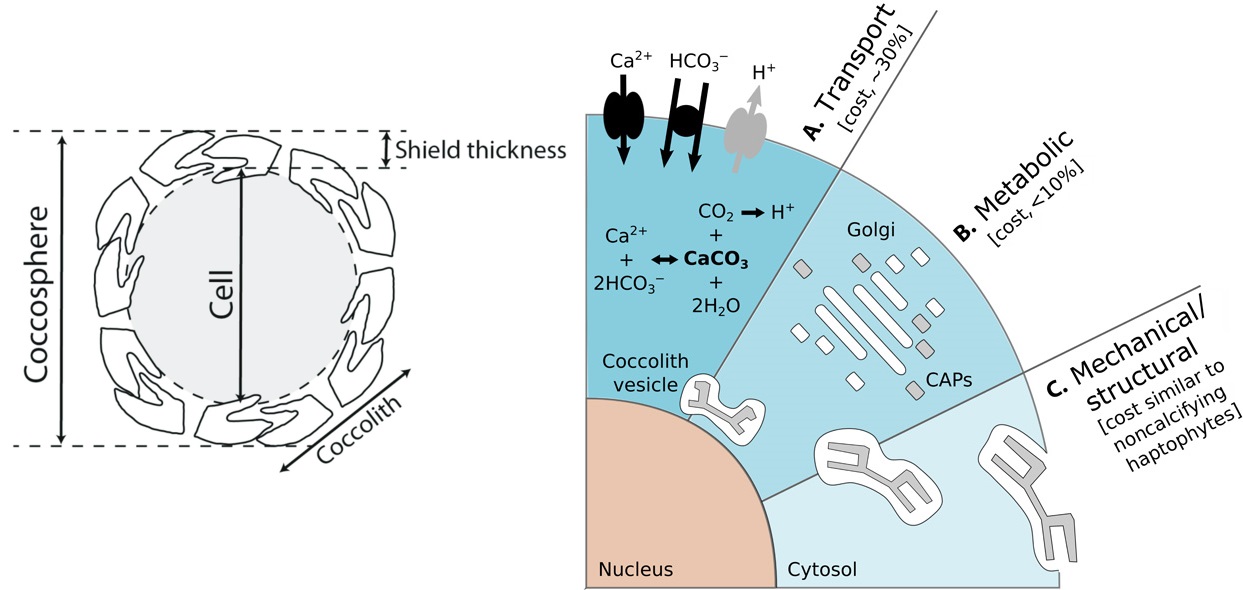
(上图:钙鞭藻细胞外铠甲及其制造过程的粗略示意。需要说明,此处的示意图严格来说只针对它们的二倍体阶段,而最近的研究证实,钙鞭藻单倍体阶段的球石也是胞内生成,但途径有所不同。具体不再展开)
钙化的另一重坏处,便是使它们对酸度变得极其敏感,这一点我们将在后面讨论。至于钙化的好处,你可能会说是为了防御掠食者。的确,铠甲可能会让一些单细胞掠食者对它无从下口,或者是运用“垃圾食物”策略,通过让自己变得难以消化来“说服”掠食者不要吃自己。但是,$2018$ 年的一项实验却表明,钙化似乎并不能明显地提升它们在掠食者威胁下的存活率。当然,钙鞭藻起源于三叠纪,那时的掠食者和现在的掠食者也不可同日而语,所以这一假说并未被完全否定。不过这也提醒了我们,钙化或许另有目的。
一种新的假说认为,钙鞭藻的铠甲可能是它们追求更高光合效率的结果。你或许已经注意到,在它们制造 $\ce{CaCO3}$ 的反应中同时会有 $\ce{CO2}$ 的产生,而研究发现,这些 $\ce{CO2}$ 将被直接用于光合作用。这样一来,钙鞭藻就有了一条快速将 $\ce{HCO3-}$ 转化为 $\ce{CO2}$ 的路线,至于 $\ce{CaCO3}$,那不过是个副产物罢了,后续能发挥什么作用纯粹是意外之喜。还有研究发现,它们的铠甲可以对光线进行许多“微操”,比如将强光的能量分散为好几份便于利用,或是像防晒衣那样遮蔽紫外线,防止其损伤细胞。此外,还有假说认为铠甲可以保护它们免受一些细菌和病毒的侵袭,等等。
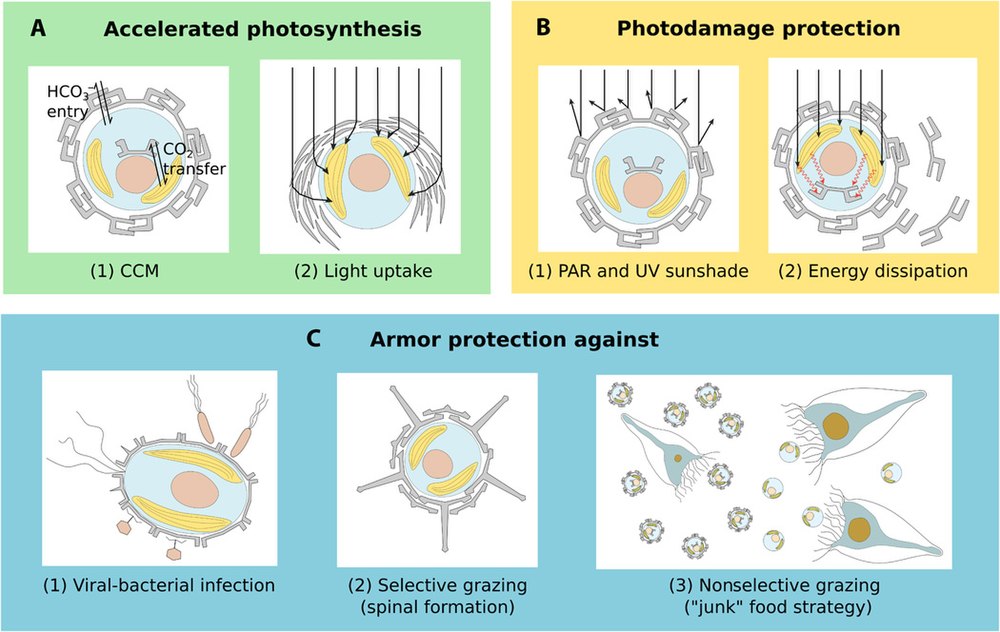
(上图:对钙鞭藻铠甲作用的几种猜测总结)
总之,铠甲一定给予了它们某些生存优势,让它们能在一众强敌之中脱颖而出。从化石记录中可以看到,钙鞭藻在约 $2.2$ 亿年前的晚三叠纪出现,随后逐渐崛起。它们最早的鳞片化石呈环形,中间有 $1 - 2$ 根轴连接,就像是车轮或者方向盘。比如生活在 $2.18$ 亿年前的古盤藻($Archaeozygodiscus$)和十字花石藻($Crucirhabdus$),被认为是这个家族最古老的类群:
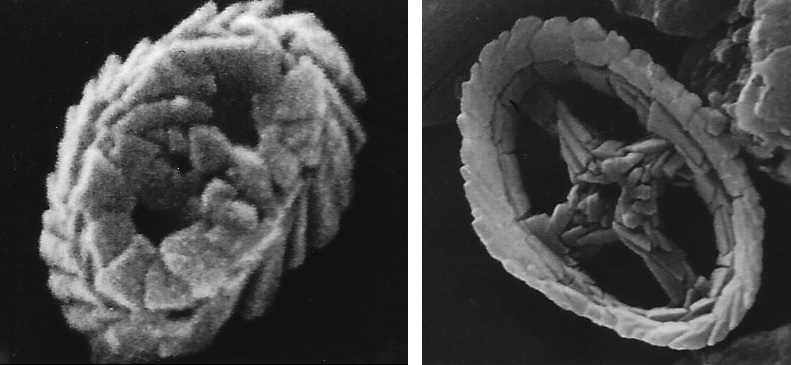
(上图:$A.~koessenensis$ (左)与 $C.~primulus$ (右)的鳞片化石。二者分别属于伊氏石藻目($\text{Eiffellithales}$)和冠石藻目($\text{Stephanolithiales}$))
在距今约 $2$ 亿年前,钙鞭藻的多样性迎来了一轮爆发,这很可能是因为三叠纪末大灭绝($\text{end-Triassic mass extinction}$)帮它打击了一些竞争对手。从侏罗纪开始,以盾石藻目($\text{Podorhabdales}$)和瓦氏石藻目($\text{Watznaueriales}$)为代表的身披厚重铠甲的类群开始逐渐占据优势。就如同古代的重装武士一般,它们的鳞片越来越厚,表面的孔隙越来越小,甚至还演化出了内外双层的复杂结构。这点从它们的化石,或者从瓦氏石藻目如今最后的孑遗——螺板藻($Tergestiella$)身上就能看出来:
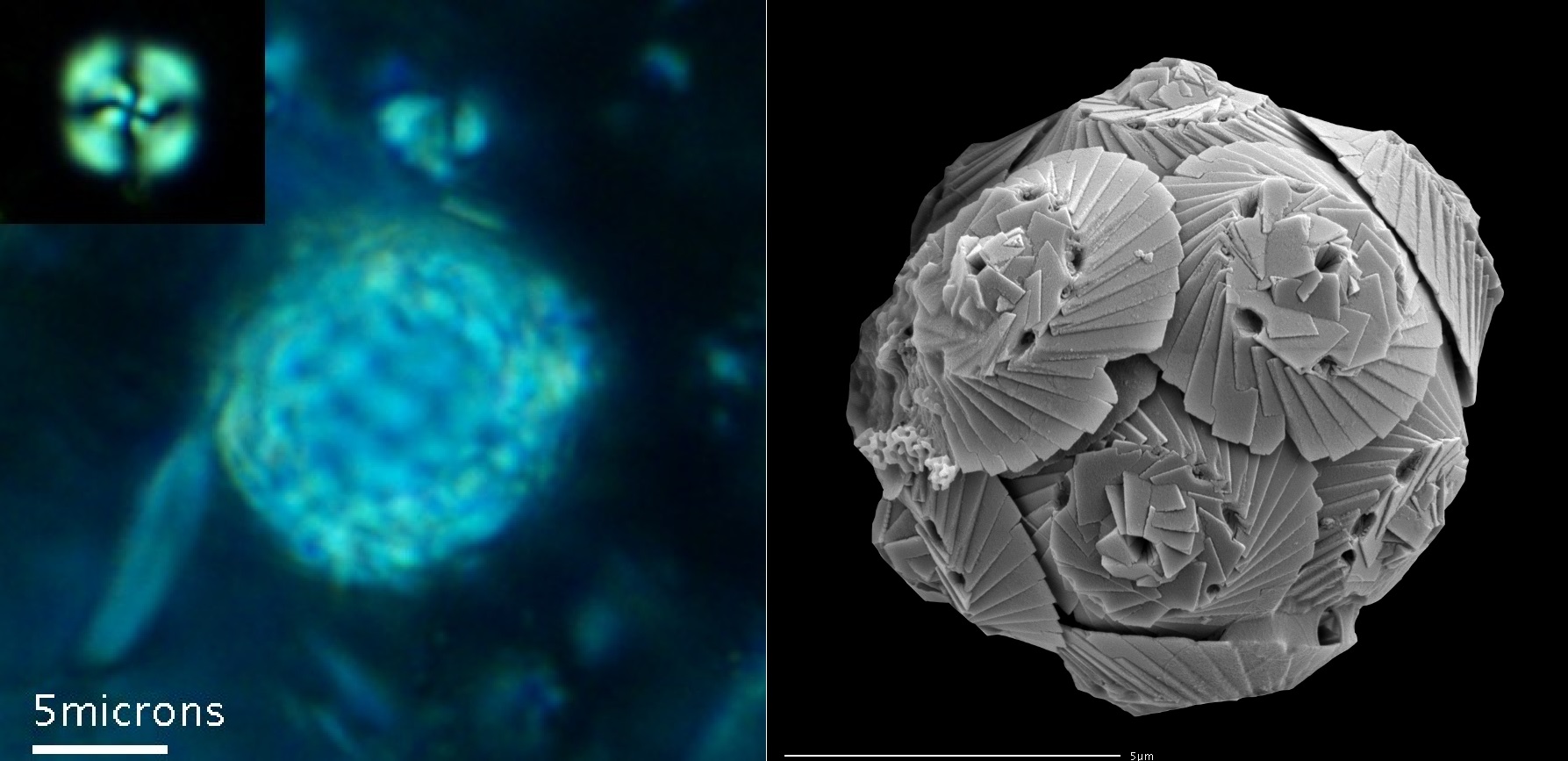
(上图:$T.~adriatica$ 的荧光照片(左)与外壳细节特写(右))

(上图:鳞片化石。第一排:瓦氏石藻目,从左到右:$Cyclagelosphaera$,$Watznaueria$,$Perchnielsenella$;第二排:盾石藻目,从左到右:$Dodekapodorhabdus$,$Miravetesina$,$Sollasites$)
到了侏罗纪晚期,随着钙鞭藻的一路高歌猛进,球石藻纲成功在浮游藻类中站稳了优势地位。但是,当时至少还有一支藻类能与它们分庭抗礼,那便是甲藻(甲藻纲,$\text{Dinophyceae}$)。说来也真巧,部分甲藻体内的质体就是来源于定鞭藻;而或许也正是因此,钙鞭藻下定了决心,要带领自己的族群打败甲藻,登上王者的宝座。
在 $1.43$ 亿年前,地球进入了白垩纪,也正是在这段时间,另外两类影响深远的钙鞭藻出现在了地层之中。其中之一是布拉鲁藻目($\text{Braarudosphaerales}$),它们对钙鞭藻传统的鳞片制造工艺进行了革新,这使得它们的外形在几何上有明显的特征。比如,从白垩纪早期存活至今的布拉鲁藻属($Braarudosphaera$),具有标志性的正五边形鳞片,有些物种($B.~bigelowii$)甚至干脆直接把自己包成了个正十二面体:

(上图:$B.~bigelowii$ 的照片(左)与外壳细节特写(右))

(上图:$B.~bigelowii$ 单个鳞片的细微结构)
其他鳞片化石也是各有各的特色。$Micrantholithus$ 属同样是 $5-$辐射对称,但它直接把鳞片干成了个五角星;$Hexadelus$ 属留下了正三角形的奇特鳞片,这些鳞片组成的铠甲则可能是个正二十面体;$Kokia$ 属某些物种的鳞片是个七角星;$Tegulalithus$ 属的鳞片非常厚,中间有漏斗状的空腔,等等:

(上图:布拉鲁藻目的鳞片化石。第一排从左到右:$Micrantholithus$ 属、$Hexadelus$ 属、$Uniplanarius$ 属;第二排:$Tegulalithus$ 属,正视与侧视图;右下:$Eprolithus$ 属;第三排从左到右:$Lithastrinus$ 属(简笔与实物)、$Kokia$ 属)
更离谱的是 $Quadrum$ 属,简直堪称 $\text{Windows}$ 的代言人。注意它的颜色,这可不是后期加上去的,而是在特定光照下由其自身的细微结构形成。我敢打赌,这可能是自然之美与人类图标设计达成的最奇特的巧合:

(上图:$Q.~gartneri$ 鳞片化石的简笔画、彩色照片与黑白照片)
虽然布拉鲁藻目从长相上看简直是奇形种,但实力的确不俗。作为晚辈,它们在白垩纪前中期飞速辐射演化,不仅在群雄林立中迅速站稳了脚跟,甚至还大有后浪推前浪,成为钙鞭藻家族新一代扛把子之势。这到底是如何做到的呢?$2024$ 年,一项新研究终于发现了它们的独门秘籍。简而言之,人们在 $B.~bigelowii$(就是上文中那个长成正十二面体的家伙)体内发现了一种前所未见的细胞器,被命名为硝质体($\text{nitroplast}$)。这种细胞器的真身是一种内共生的蓝细菌(蓝菌门,$\text{Cyanobacteria}$),它允许 $B.~bigelowii$ 直接进行固氮,即利用空气中的氮气来制造一些生命活动所需的物质:
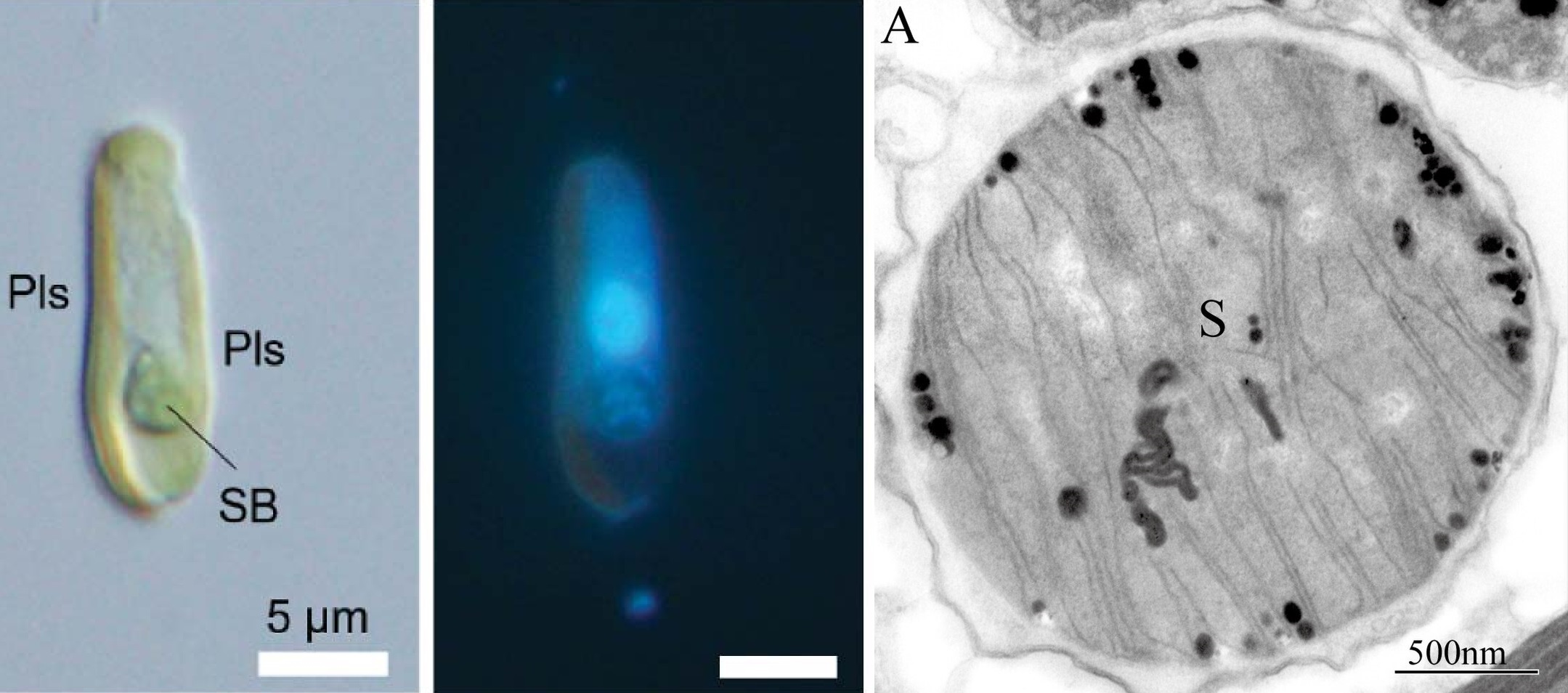
(上图左中:人为去掉外壳后的 $B.~bigelowii$ 的普通照片与荧光照片;右:硝质体特写,透射电镜照片;$\text{SB}$ 或 $\text{S}$:硝质体)
作为人类发现的第一种真正掌握了固氮作用的真核生物,这种开挂般的配置不仅改写了教科书,也在很大程度上改写了这个族群的命运。据估计,这次内共生发生在一亿多年前,与布拉鲁藻目辐射演化的时间点基本一致。因此,它们之所以能够脱颖而出,很有可能就是因为有了硝质体。
在 $1.1$ 亿年前的中白垩纪,地球迎来了一波急剧的全球变暖,史称帕奎尔-乌尔比诺极热事件($\text{Paquier/Urbino Thermal Maximum}$),此后的两千多万年,地球气温总体上都居高不下,史称中白垩纪温室期($\text{Mid-Cretaceous Hothouse}$)。这段时期,在高温以及爆表的 $\ce{CO2}$ 浓度的加持下,整个球石藻纲迎来了演化史上最大的爆发。此后虽然一头撞上了 $9400$ 万年前的森诺曼期-土仑期灭绝事件($\text{Cenomanian-Turonian extinction}$,下图 $\text{OAE-2}$),但也无伤大雅。在晚白垩纪,球石藻的多样性持续上升,至 $7500$ 万年前左右达到巅峰。这段时间,从淡水到海洋,球石藻的数量多到满坑满谷,它们也终于如愿以偿地拿到了藻类之王的皇冠。
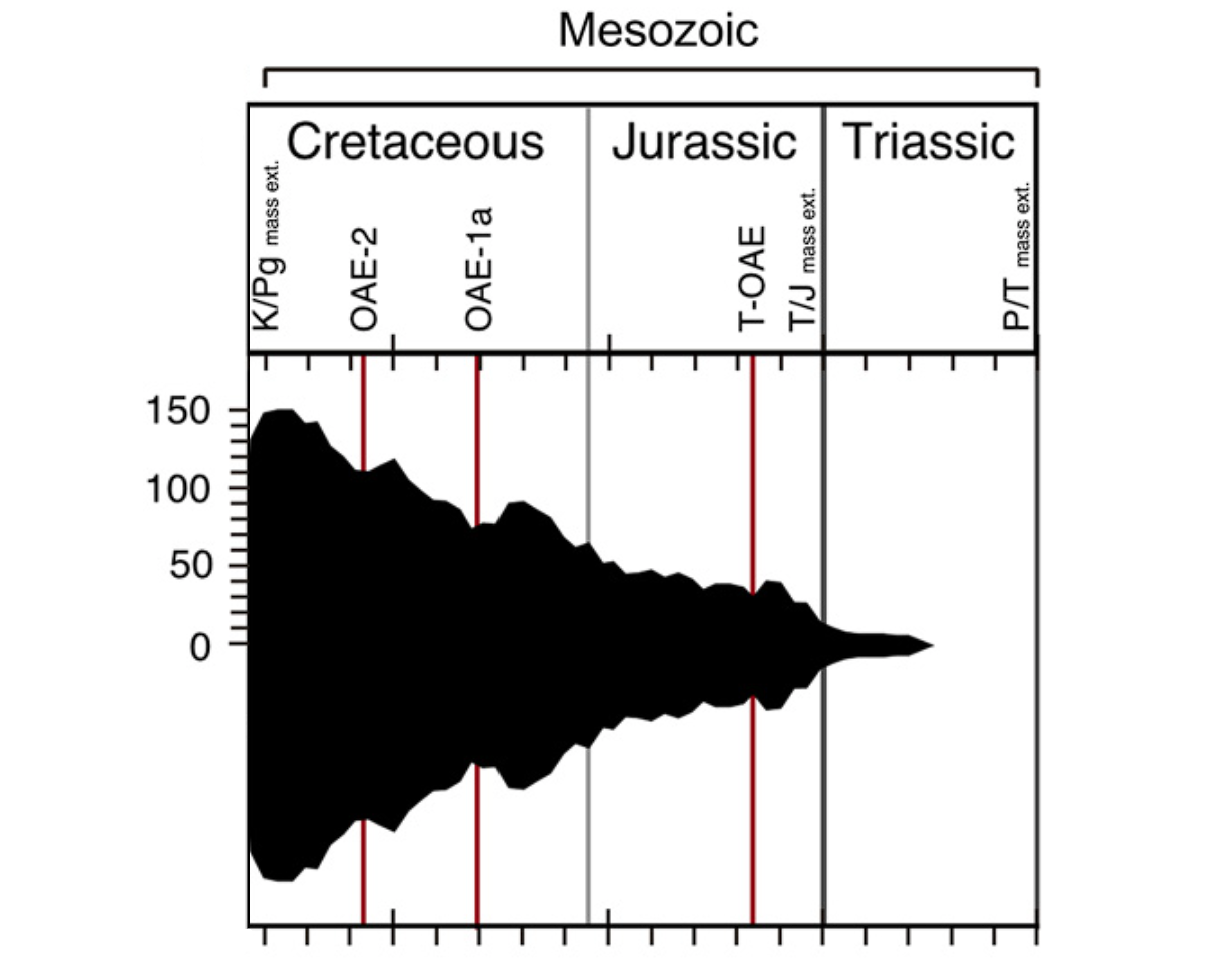
(上图:中生代球石藻种类丰富程度随时间的变化)
只可惜,当球石藻达到极盛之日,整个中生代的余额却已不足一千万年;而它们也终究无法预料,那无常命运的戏弄。
距今 $6600$ 万年前,一颗迷途的大陨石让地球的生物圈直接重开,史称白垩纪末大灭绝($\text{end-Cretaceous mass extinction}$)。事实上在灭绝的前夕,球石藻的多样性已经开始下降(从上图也可以看到),可能是因为更早发生的德干暗色岩事件($\text{Deccan Traps}$)已经拉开了海洋酸化的序幕。在如此巨大的天灾之下,整个定鞭藻门 $90\%$ 以上的物种直接绝灭,而作为中流砥柱的钙鞭藻亚纲更是遭遇毁灭性打击。我们可以来盘点一下钙鞭藻各类群的损失:
\[\ce{CaCO3 + 2H+ -> Ca^{2+} + H2O + CO2}\]阿尔汉石藻目($\text{Arkhangelskiales}$)全部灭绝;
布拉鲁藻目($\text{Braarudosphaerales}$)仅剩两属,但如今仅存一属两种;
伊氏石藻目($\text{Eiffellithales}$)全部灭绝;
微棒石藻目($\text{Microrhabdulales}$)全部灭绝;
盾石藻目($\text{Podorhabdales}$)仅剩两属,但都在古近纪中期灭绝;
冠石藻目($\text{Stephanolithiales}$)全部灭绝;
瓦氏石藻目($\text{Watznaueriales}$)仅剩三属,又有两属在古近纪早期灭绝,实际存活仅一属一种;
其他还有许多未能归入任何目的钙鞭藻,基本都未能挺过大灭绝。
(注:这其实并非钙鞭藻亚纲的全部,为防剧透,部分类群被刻意隐去)
很多人都直接拿上面这个方程式来解释为什么它们在大灭绝中遭受了如此致命的打击,但事实上,这有些过于简化了。的确,陨石撞击等一系列事件导致了严重的海水酸化,但对钙鞭藻来说真正的麻烦在于,它们制造铠甲的过程中需要向外排出氢离子。现在是时候回收之前埋下的伏笔了,让我们把这张图再放一遍:
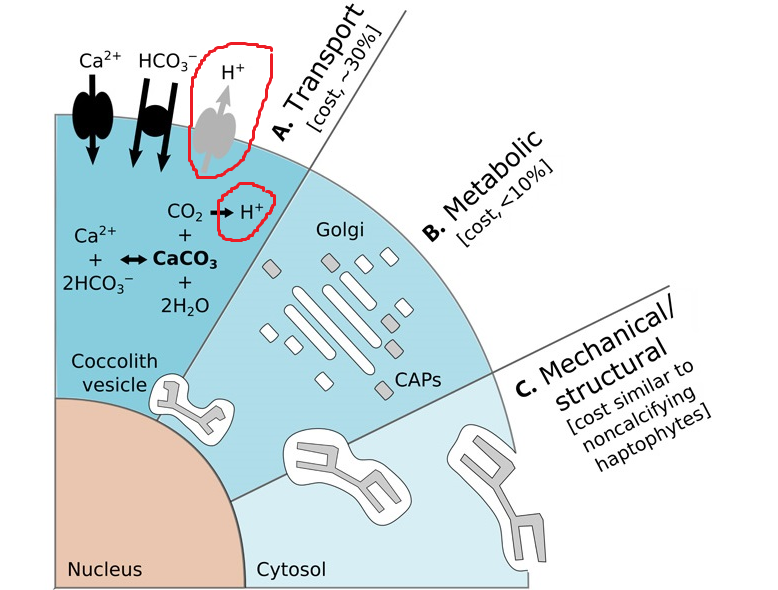
研究表明,钙鞭藻泵入胞内的 $\ce{Ca^{2+}}$ 和 $\ce{HCO3-}$ 实际比例达不到 $1:2$,$\ce{Ca^{2+}}$ 总是更多一些,如此便会导致多余的正电荷,并最终以 $\ce{H+}$ 的形式排出。随着外界环境 $\text{pH}$ 值的降低,细胞内多余的 $\ce{H+}$ 会越来越难以排出,直至耗尽细胞内的缓冲机制。此时,一系列对酸性非常敏感的细胞器将无法正常运作,最终导致整个细胞的崩溃。或许直到此刻,它们才如梦初醒,但却为时已晚——在此前两亿多年的演化中,那越来越精巧、复杂、厚重的铠甲早已无法脱去,它们便只能眼睁睁地看着自己,与那曾经引以为傲的铠甲一起坠入深渊。无言的大海并没有挽留它们,只是不断地搬运这它们的遗骸,最终形成了白垩,形成了今天震撼人心的白色坟场。

(上图:白垩。在引言中没有讲到,白垩常被用来制作粉笔,或许你曾经在黑板上书写的字迹中,就有千万年前这些小藻的微微残躯)
那么,难道钙鞭藻真的就这样离我们远去了吗?这位中生代的藻类王者,真的就只剩下这么几支后裔在苟延残喘了吗?
后章:新生
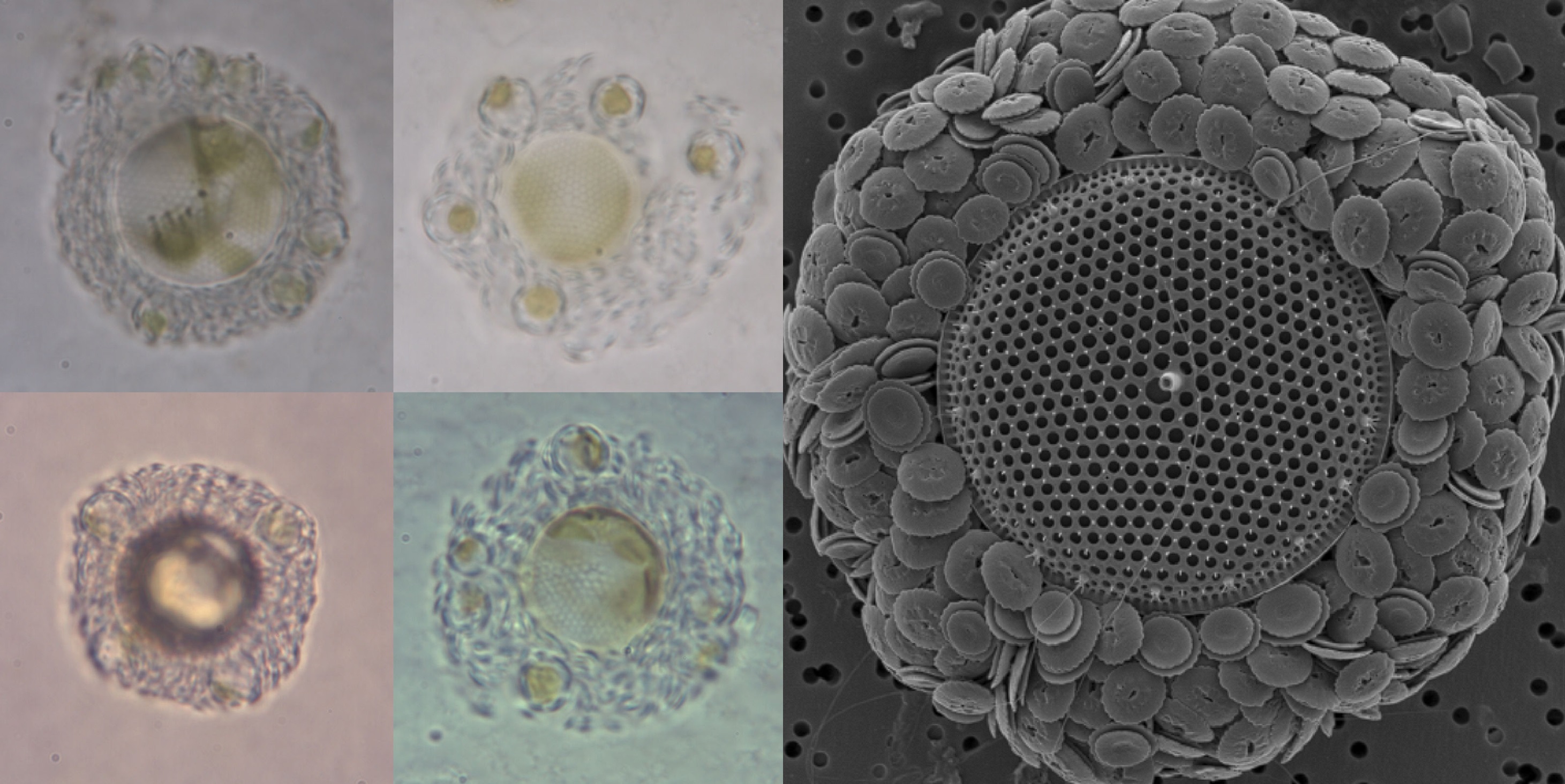
我们需要把进度条拉回白垩纪早期。在布拉鲁藻出道即横扫四海之日,另一类影响深远的钙鞭藻也出现了,那便是条结球藻目($\text{Syracosphaerales}$)。只是在那会,它们还是一个被排挤到边缘的小派系,有些研究认为它们可能是冠石藻目中的一支,且很可能与冠石藻属($Stradnerlithus$)紧密相关,但也未能确定。不过,它们确实在演化方向上与其他钙鞭藻有很大不同。
最鲜明的一点,便是它们没有追求更大的体型,也没有演化出更加复杂厚重的鳞片,只是在最原始的“方向盘”上略做更改,添置了更多轴来连接,从而使其兼顾轻便与强度:
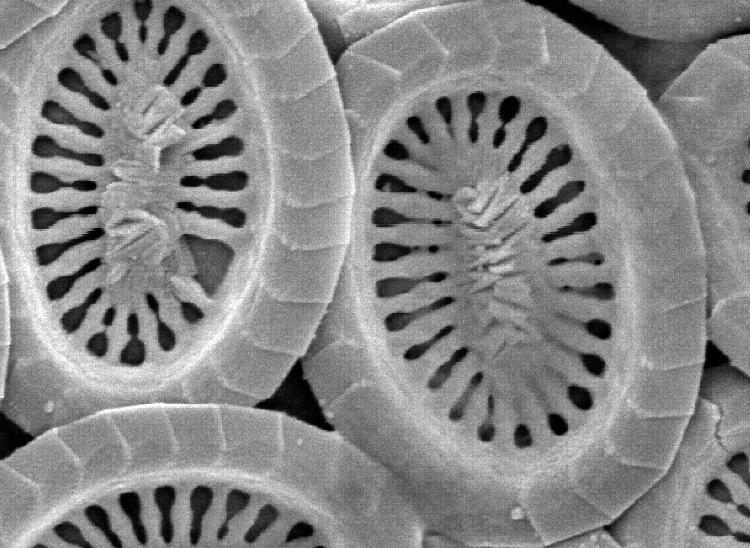
(上图:典型的条结球藻鳞片形态)
或许是因为生存空间的逼仄,有一支早期的条结球藻刚出门就立马拐上了邪道。它们把自己的身体拉成了长条,覆盖于其上的鳞片也从椭圆变成了菱形,四条边向上突起,乍一看就跟个小船似的,因此得名舟管藻($Calciosolenia$)。不过,这艘“小船”是漏水的,因为其船底还保留了祖传的轴结构,而并非平整的一块:

(上图:$C.~corsellii$ 及其鳞片特写。附注:舟管藻属约在一亿三千多万年前分化,存活至今)
当然,大多数条结球藻还是规规矩矩地保持着原有的形态,在整个白垩纪,它们都以微小的身躯和稀少的数量,游走于其他强势类群的狭缝中,聊以求生。这段时间,它们留下的仅是一些破碎的鳞片化石,多样性也始终停滞在属级别:
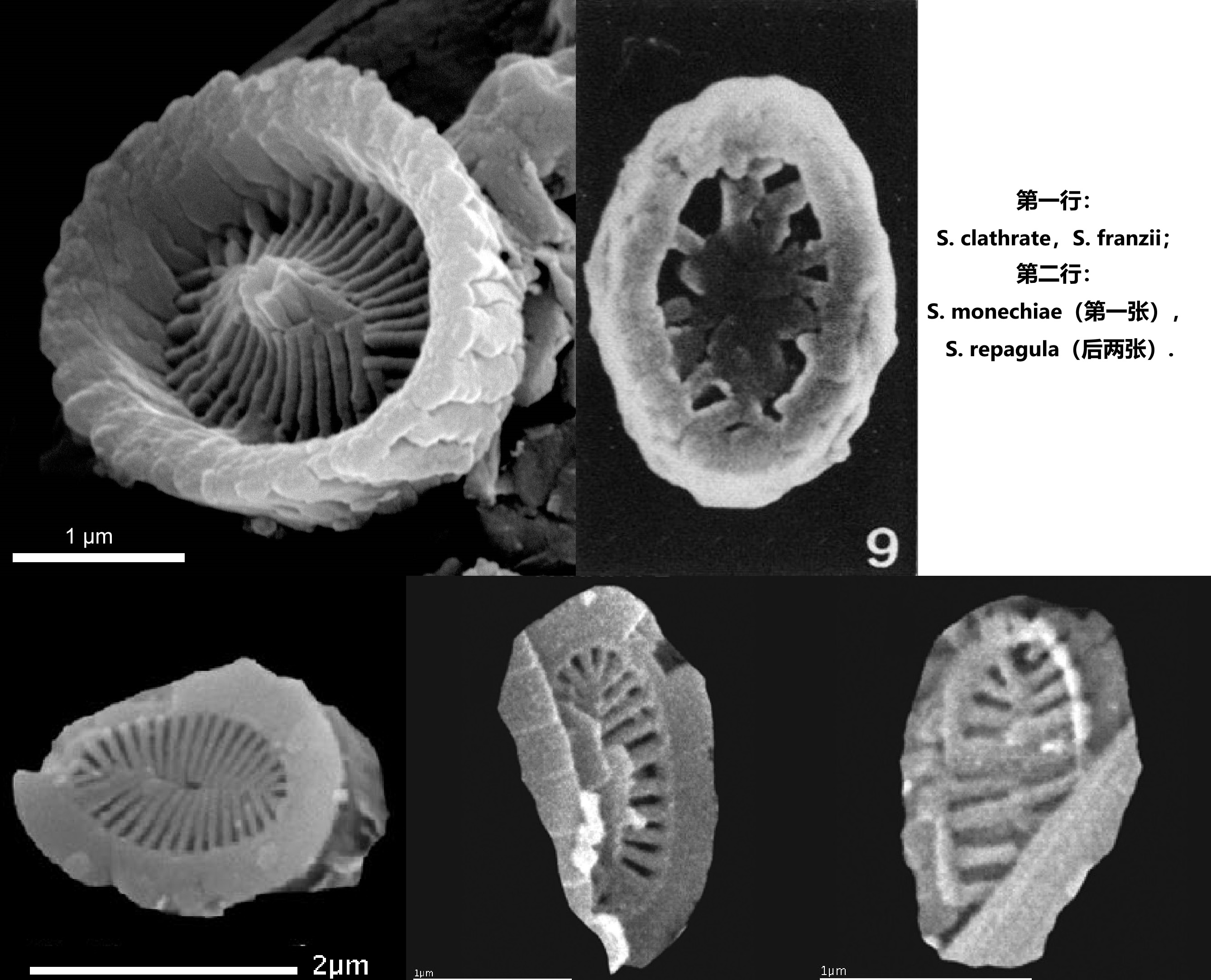
(上图:条结球藻鳞片化石)
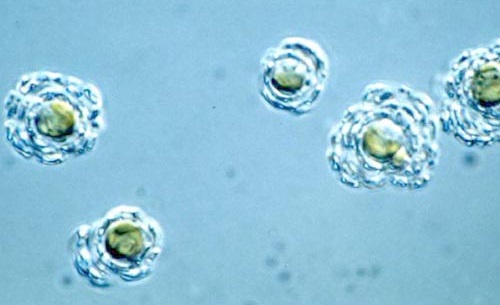
或许是因为无力与其他钙鞭藻争夺钙质,许多条结球藻都在反向的道路上越走越远,将自己曾引以为傲的铠甲打薄,还在上头各种镂空、雕花,以减少对钙的需求。这股逆流终于在白垩纪晚期达到顶峰——有一支条结球藻彻底放弃了单倍体阶段的钙质铠甲,仅保留一些有机鳞片或者干脆就裸奔。它们演化为了如今的等鞭金藻目($\text{Isochrysidales}$)。
为了更好地理解它们这违背祖宗的决定,我们先得解释一下球石藻的世代交替。众所周知,人类的体细胞是二倍体,生殖细胞是单倍体,而球石藻虽然(绝大多数)是单细胞生物,但同样有此区分。不同之处在于,人类的单倍体生殖细胞不可能独立存活,但球石藻的单倍体和二倍体不但都能独立生活,而且还有不同的形态。最重要的一点是,单倍体细胞和双倍体细胞都可以无性繁殖,也可以相互转化,具体如下图所示:
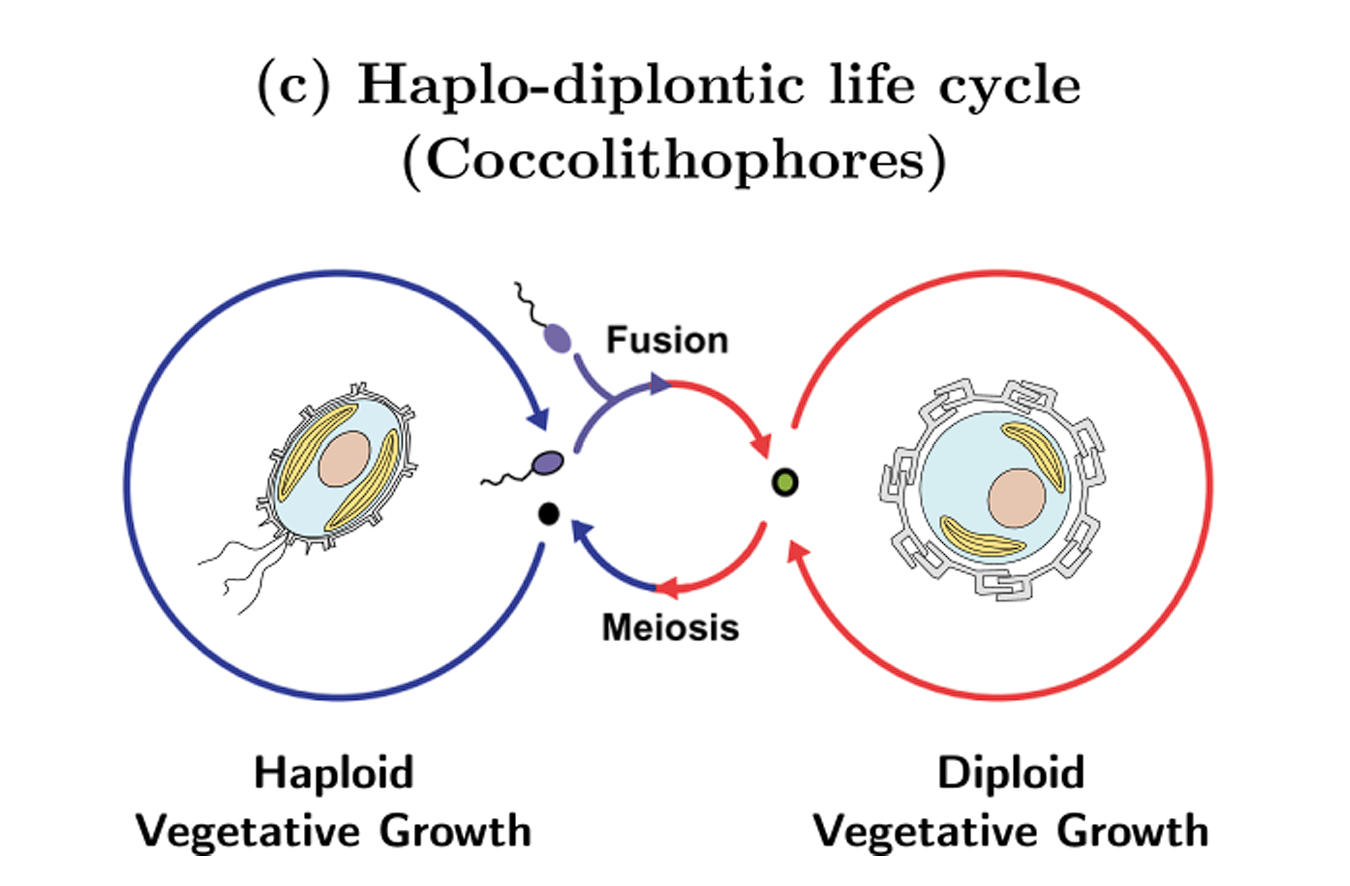
这样一来,就可以理解为何等鞭金藻目能够以相对较小的损失熬过大灭绝了。当海水酸化之时,因为它们的单倍体细胞没有钙化,所以不会受到太大的影响;又因为单倍体细胞可以独立生活,也能无性繁殖,所以整个种群得以维持,不至于崩溃。而通过了大灭绝的筛选之后,等鞭金藻目也顺理成章地成为了新生代最繁盛的钙鞭藻类群之一。
不过,它们中的一些似乎也有点上头了。可能在新生代早期,就有一支等鞭金藻彻底转向了装甲无用论,便一不做二不休,把二倍体阶段的钙质铠甲也给退化掉了,向老前辈普林藻目看齐。它们便是等鞭金藻科($\text{Isochrysidaceae}$),钙鞭藻中最大逆不道的类群。但不得不承认,它们直到今日依然繁盛,而且海洋淡水两开花;部分种类还被人类看中,成为了水产养殖业的重要饲料:
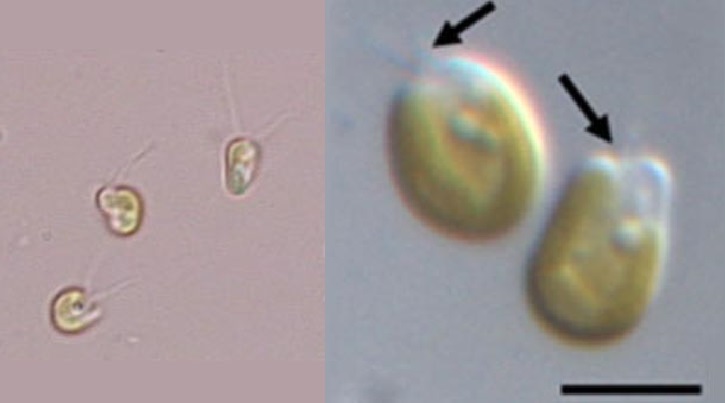
(上图:等鞭金藻属的 $I.~galbana$ 和异等鞭金藻属的 $T.~lutea$,二者都是重要的水产饲料,尤其是前者大量被用于喂养海生双壳类)
相比之下,柄石藻科($\text{Noelaerhabdaceae}$)就没那么激进了,始终保持着二倍体阶段的钙化。该科中最早出现的类群是窗孔藻属($Reticulofenestra$),顾名思义,它的(其中一种)鳞片上密密麻麻地布满了小孔,对密恐患者有极强的杀伤能力:
(上图:$R.~sessilis$ 的照片与鳞片特写)
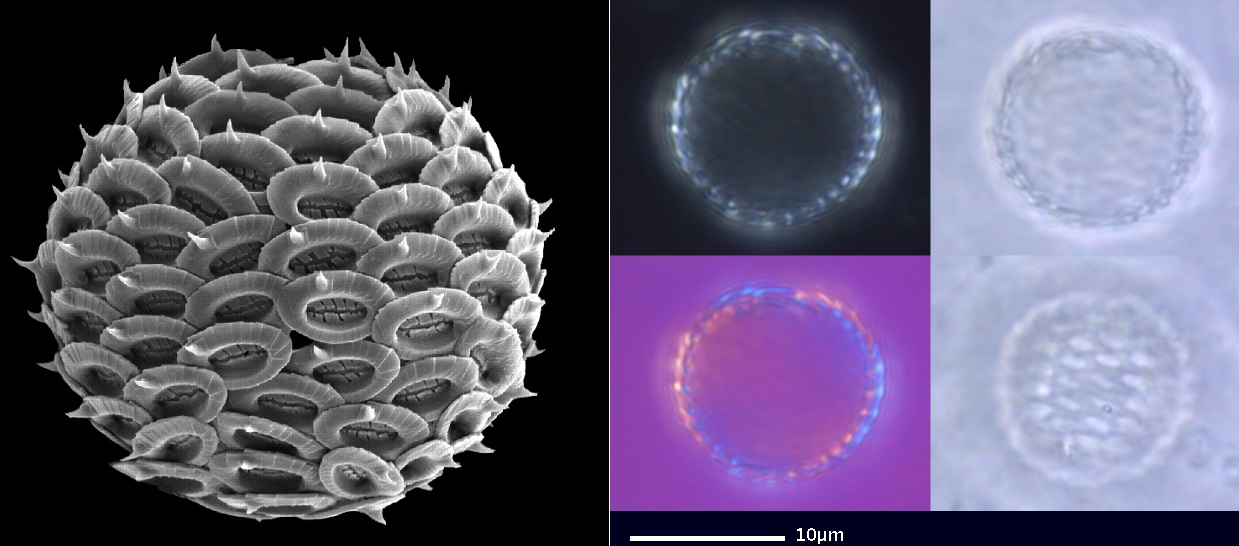
同科近亲艾氏石藻属($Emiliania$)虽然仅有 $E.~huxleyi$ 一个物种,但却是如今数量最多的球石藻(也是体型最小的球石藻之一)。从赤道到极地,其身影遍布世界每一处海洋,在某些地区甚至能以一己之力占到球石藻整体数量的 $40\%$ 以上。当它形成藻华之时,由于其钙质铠甲能够有效地折射光线,所以会对那片海域的反照率产生明显影响,形成俗称为“白水”($\text{white waters}$)的现象。“白水”经常覆盖上万平方公里,甚至明显到可以被太空中的卫星捕捉。
(上图:$E.~huxleyi$)

(上图:$E.~huxleyi$ 的细胞表面形态和单个鳞片手绘)

(上图:$1999$ 年英吉利海峡的 $E.~huxleyi$ 藻华)
在等鞭金藻目之外,大部分条结球藻虽没有那么极端,但也在很大程度上挺过了大灭绝。当余波过去,地球环境再度趋于平稳之日,它们便迅速接手了先辈的衣钵,重新辐射演化为一个庞杂的类群。只不过遭此一劫之后,球石藻的江山已经易手。一支名为硅藻(硅藻纲,$\text{Bacillariophyceae}$)的淡色藻抢先了一步,从此便稳坐新生代最强藻类的宝座,而球石藻又要回去和甲藻争夺老二了。
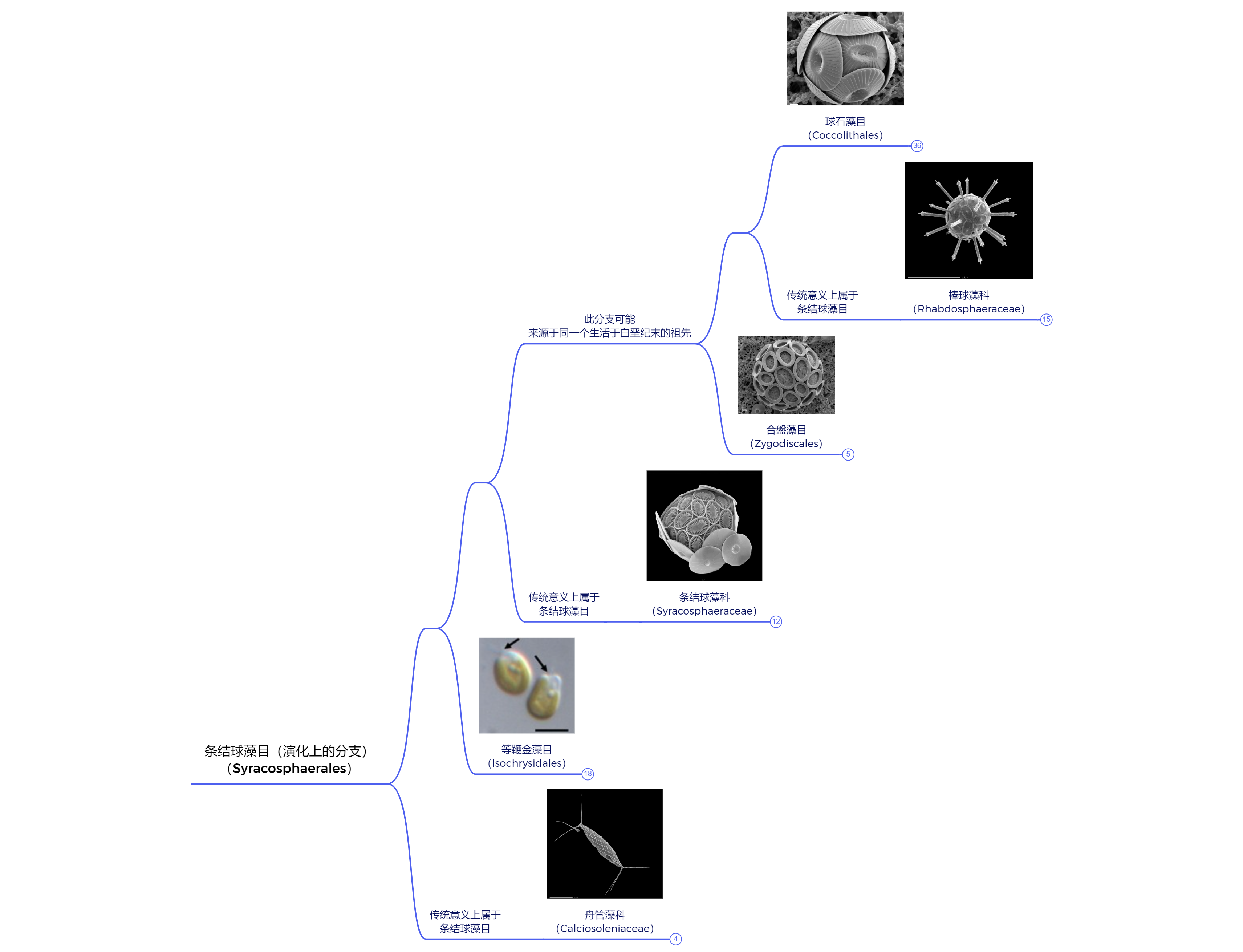
(上图:现存条结球藻各类群之间的关系)
但至少,家族中的新锐已经登上了历史舞台。上文提到,中生代那些铠甲厚重的大型钙鞭藻已经十不存一,而眼瞅着这生态位的空缺,合盤藻目($\text{Zygodiscales}$)就动起了心思。它们迅速将自己的铠甲加厚,体型增大,最终得以睥睨众藻。如今,可以长到 $50μm$ 的坛球藻($Scyphosphaera$)依旧繁盛于全球海域,而其名字来源就是铠甲外面额外长出了一圈类似老坛酸菜发酵罐的东西,被称为篮球石($\text{lopadolith}$)。据推测,该结构可能会起到引导光线的作用:
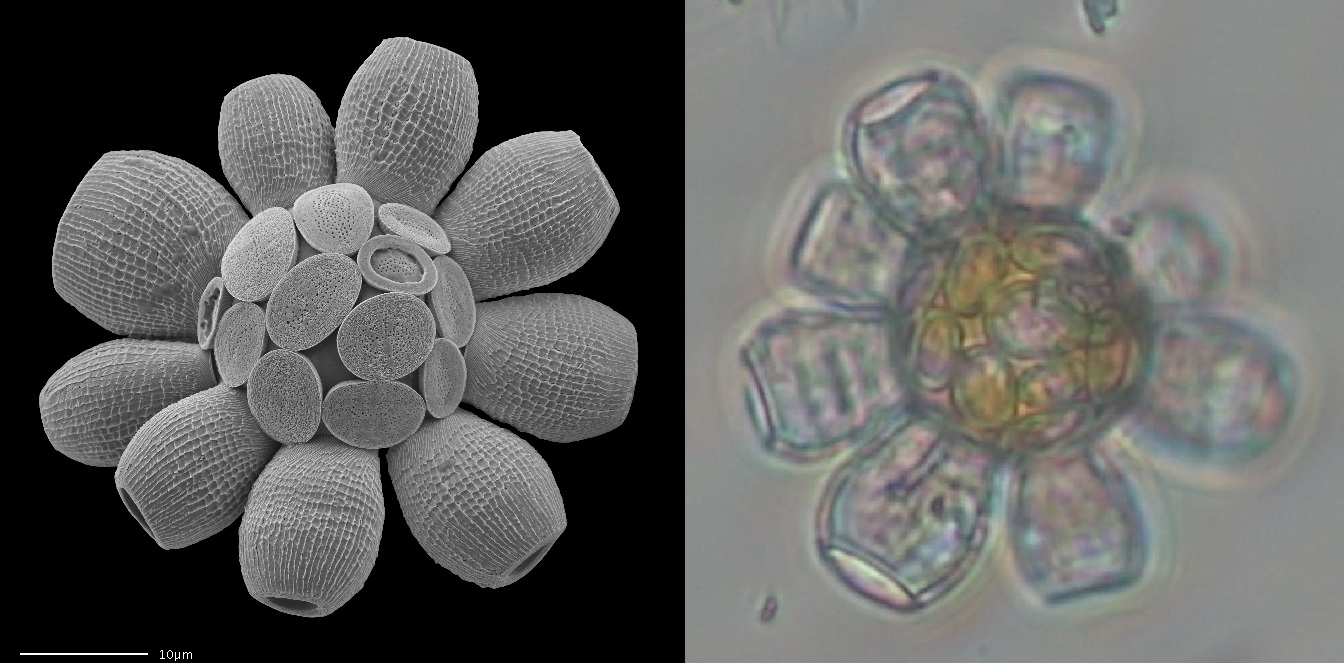
(上图:$S.~apsteinii$)

(上图:$S.~porosa$,及其篮球石特写。相比于 $S.~apsteinii$,它的篮球石更加“瘦长”)
棒球藻科($\text{Rhabdosphaeraceae}$)则换了一个思路,它们将某些鳞片的中心部位向外突出,从而改造成细长的尖刺用以防身。在一些属中,其长度甚至可以超过细胞本体的直径。冑甲球藻属($Palusphaera$)更是干脆让所有鳞片都长出尖刺,甚至还嫌不够,又给一些刺额外添上了分叉,从而使其彻底包围全身,令人见之胆寒:
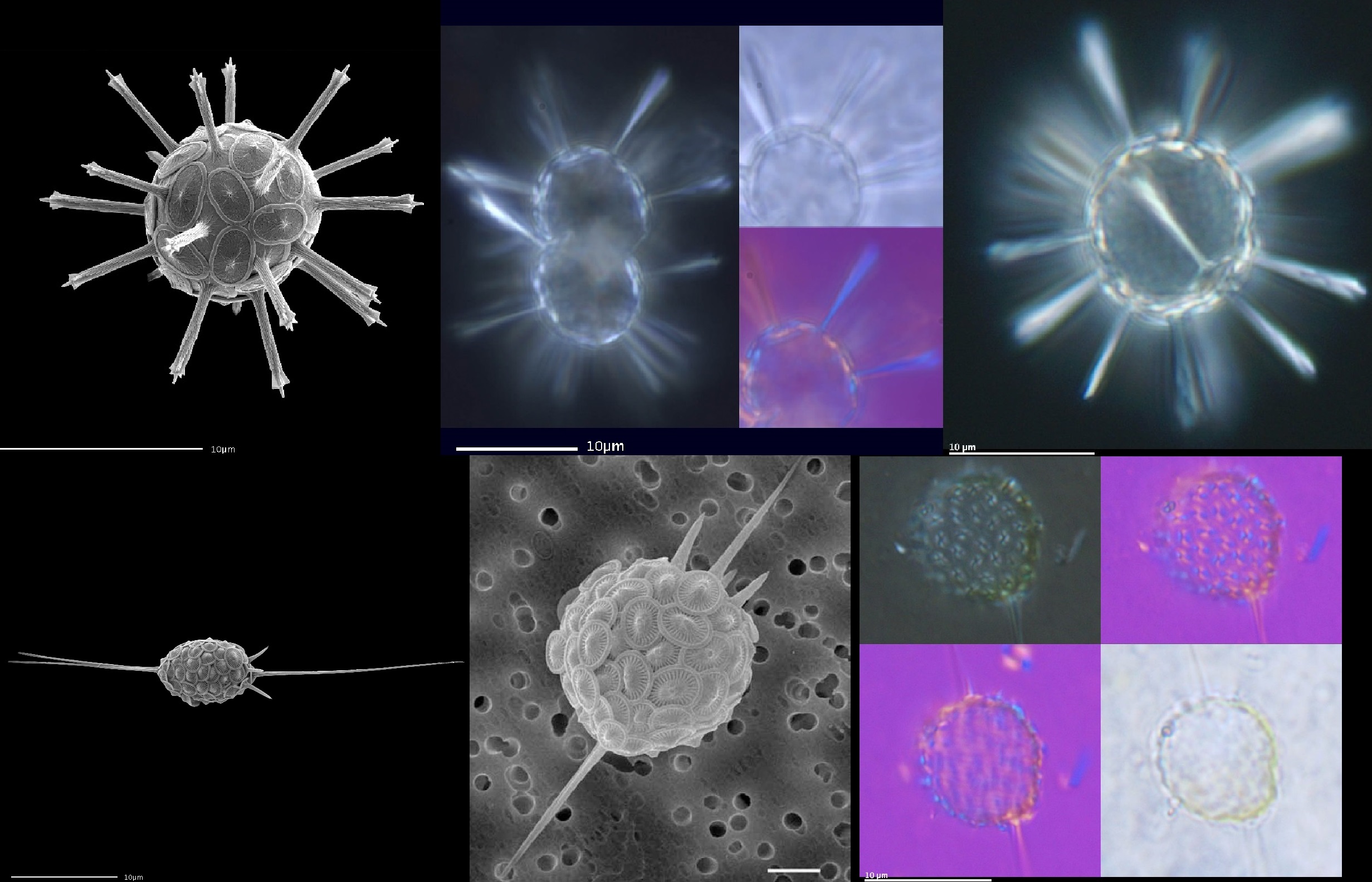
(上图:第一排:棒球藻属($Rhabdosphaera$),$R.~clavigera$;第二排:针刺藻属($Acanthoica$),$A.~quattrospina$)
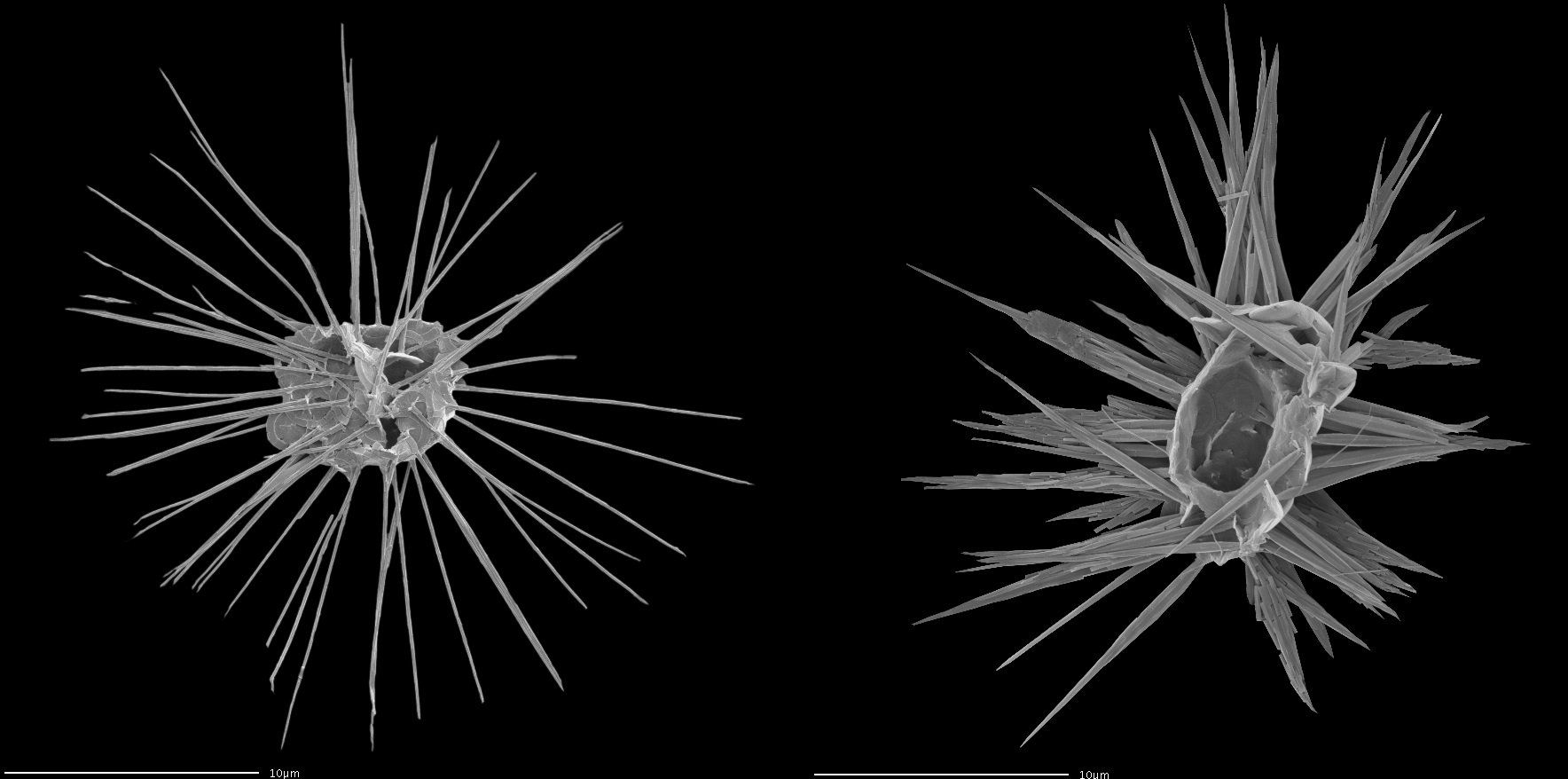
(上图:冑甲球藻属的 $P.~vandelii$(左)与 $P.~crosiae$(右))
好吧,作为一个形态多样性很高的类群,棒球藻科里也并非全都是尖刺反伤流。比如在我国近海很常见的环翼藻($Algirosphaera$),特殊的突起结构使它的鳞片看起来像顶小帽子(虽然这个尺寸估计只有细菌能戴)。更加奇怪的还有盘球藻($Discosphaera$),它的鳞片上仿佛长出了一个大喇叭,而我们至今仍不清楚这些结构究竟有什么作用:
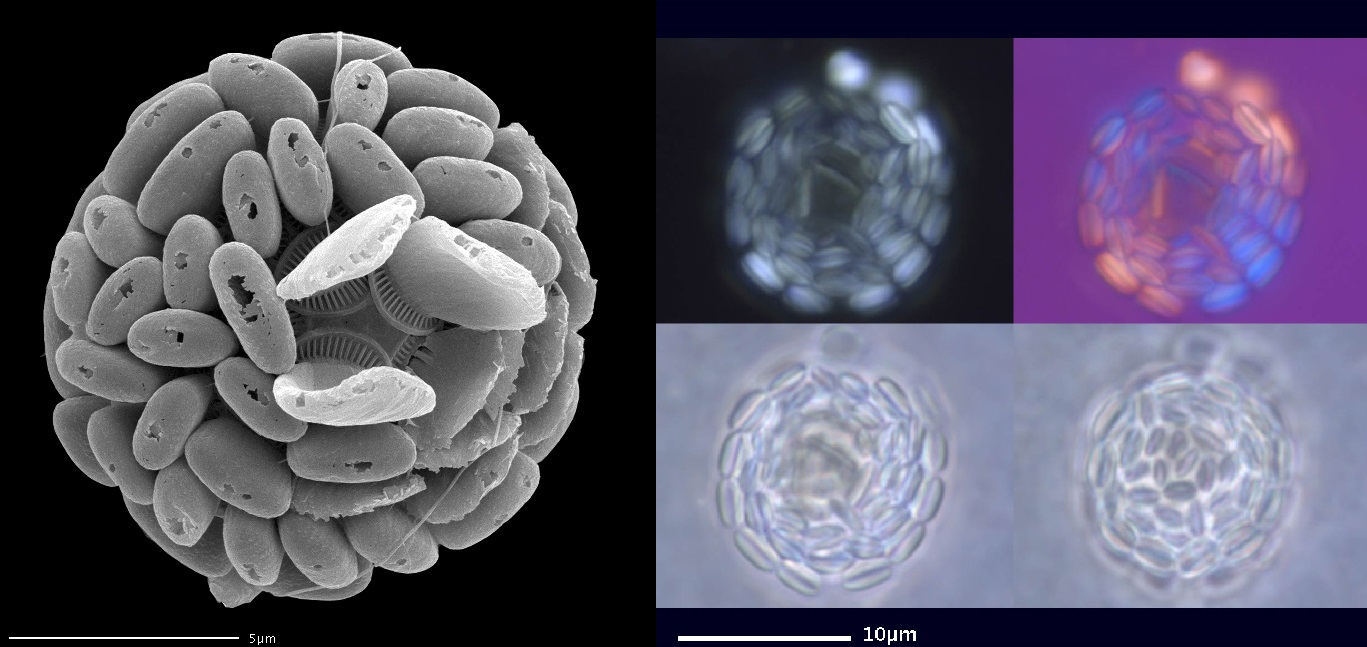
(上图:环翼藻属的 $A.~robusta$)

(上图:环翼藻的鳞片化石。左列:$A.~meteora$;右列:$A.~robusta$(上)与 $A.~cucullata$(下))
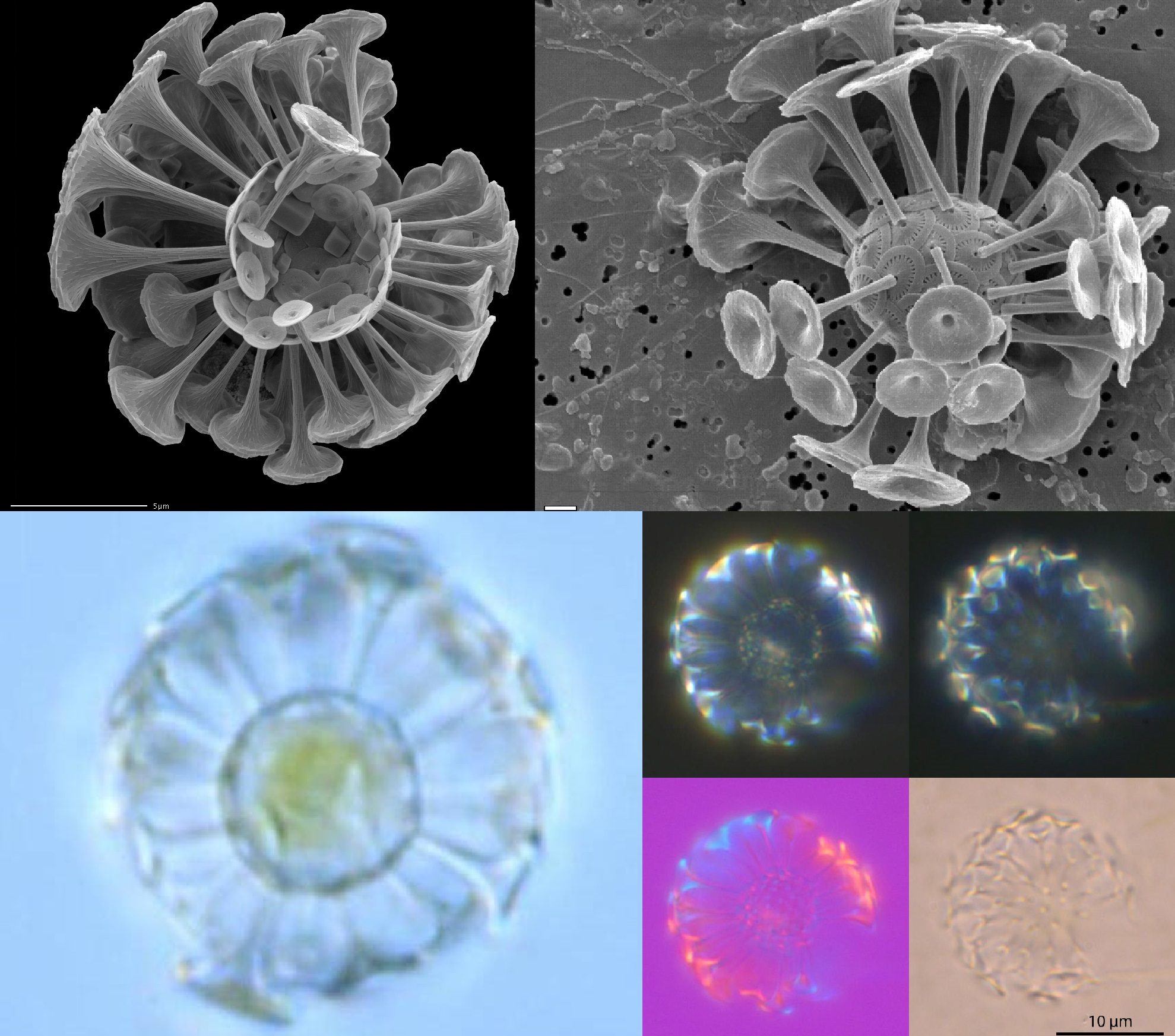
(上图:盘球藻属的 $D.~tubifera$)
当然,若真要论多样性,那球石藻目($\text{Coccolithales}$)肯定不会服气,毕竟这与纲一致的名字就足以证明其成功。该目一度拥有近 $30$ 个属(虽然现存的只剩 $10$ 个了),也包含了我们认知中最经典的那些球石藻,比如球石藻属($\text{Coccolithus}$)和钙板金藻属($\text{Calcidiscus}$)。它们的长相可以用普通来形容,既没有什么特化的奇形怪状的鳞片,也没有什么附属物,在低倍的光学显微镜下只是一个亮闪闪的球而已:
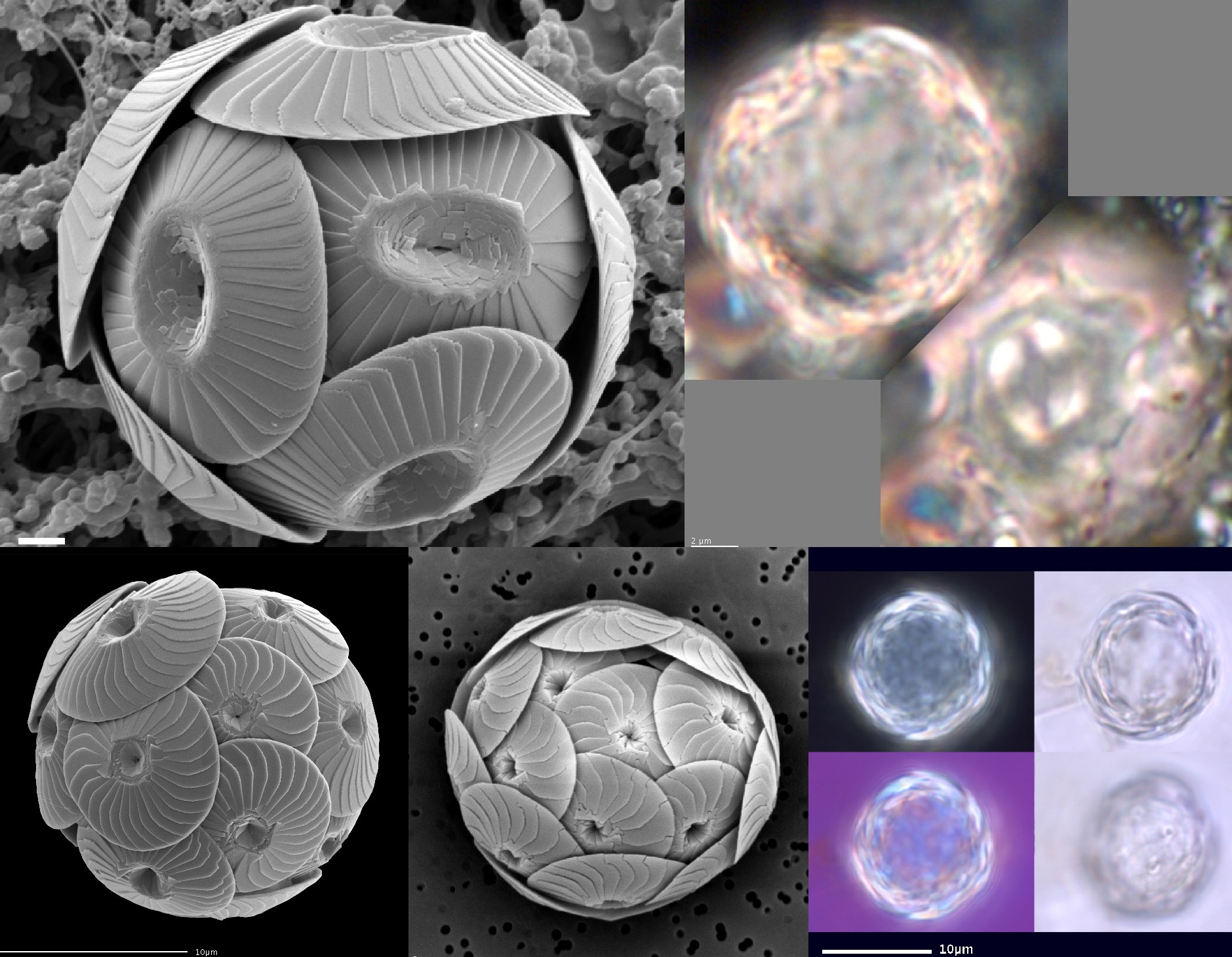
(上图:第一行:球石藻属的 $C.~pelagicus$;第二行:钙板金藻属的 $C.~leptoporus$)
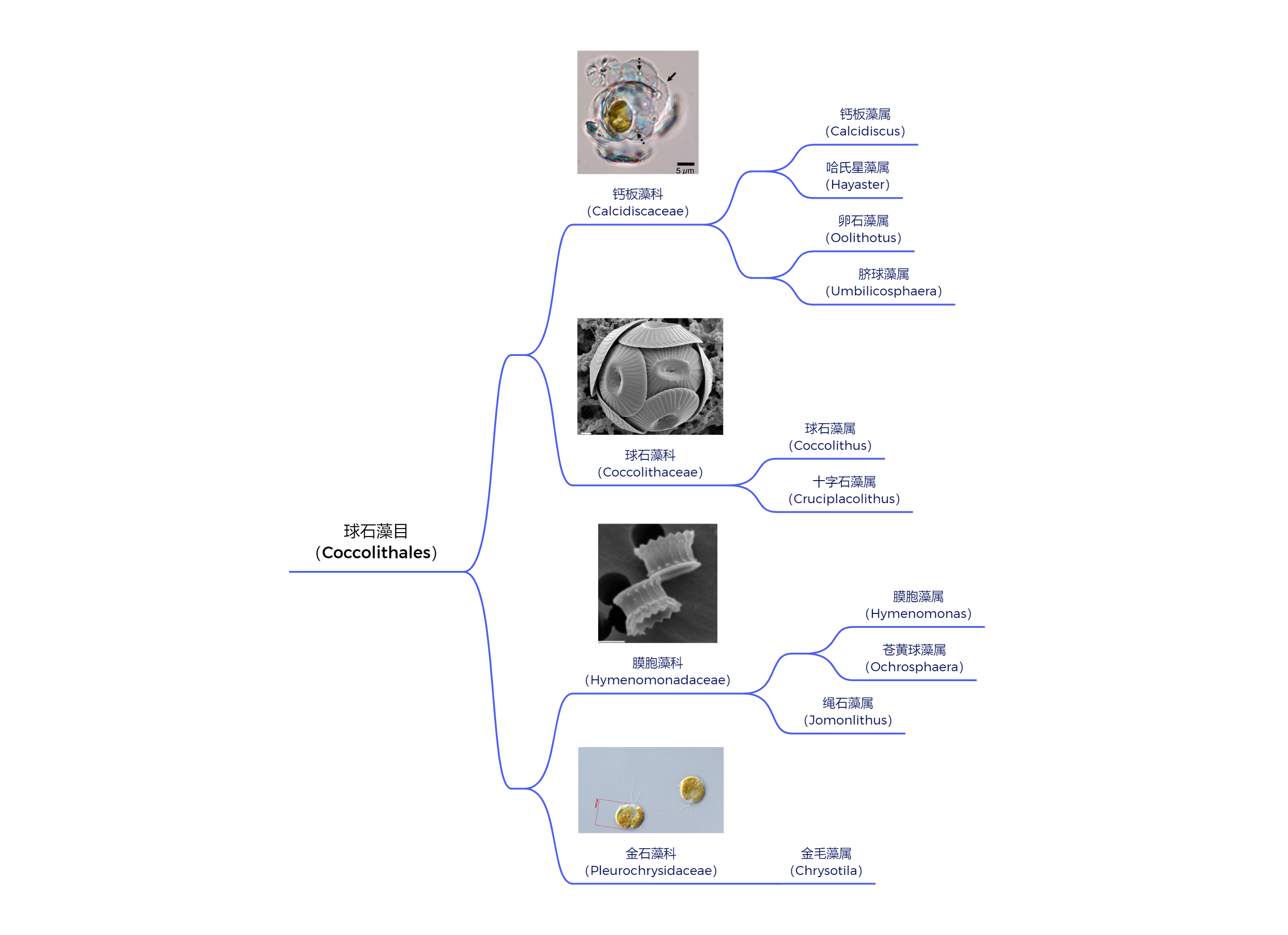
(上图:球石藻目现生类群演化树)
在距今 $5580$ 万年前,地球突然就给这些开启二周目的新玩家送上了一份大礼。只见在极短的事件内,全球平均气温突然飙升了 $5-8$ 度,史称古新世-始新世极热事件($\text{Paleocene–Eocene thermal maximum}$,$\text{PETM}$)。几乎是在一瞬之间,球石藻的多样性急剧上升(见下图),峰值甚至与白垩纪末,那盛世的最后时刻齐平。但就当所有人都认为球石藻第二帝国指日可待之时,它们的多样性却又随着极热事件的结束骤然暴跌,以至于在地质纪年下形成了一个明显的尖峰:
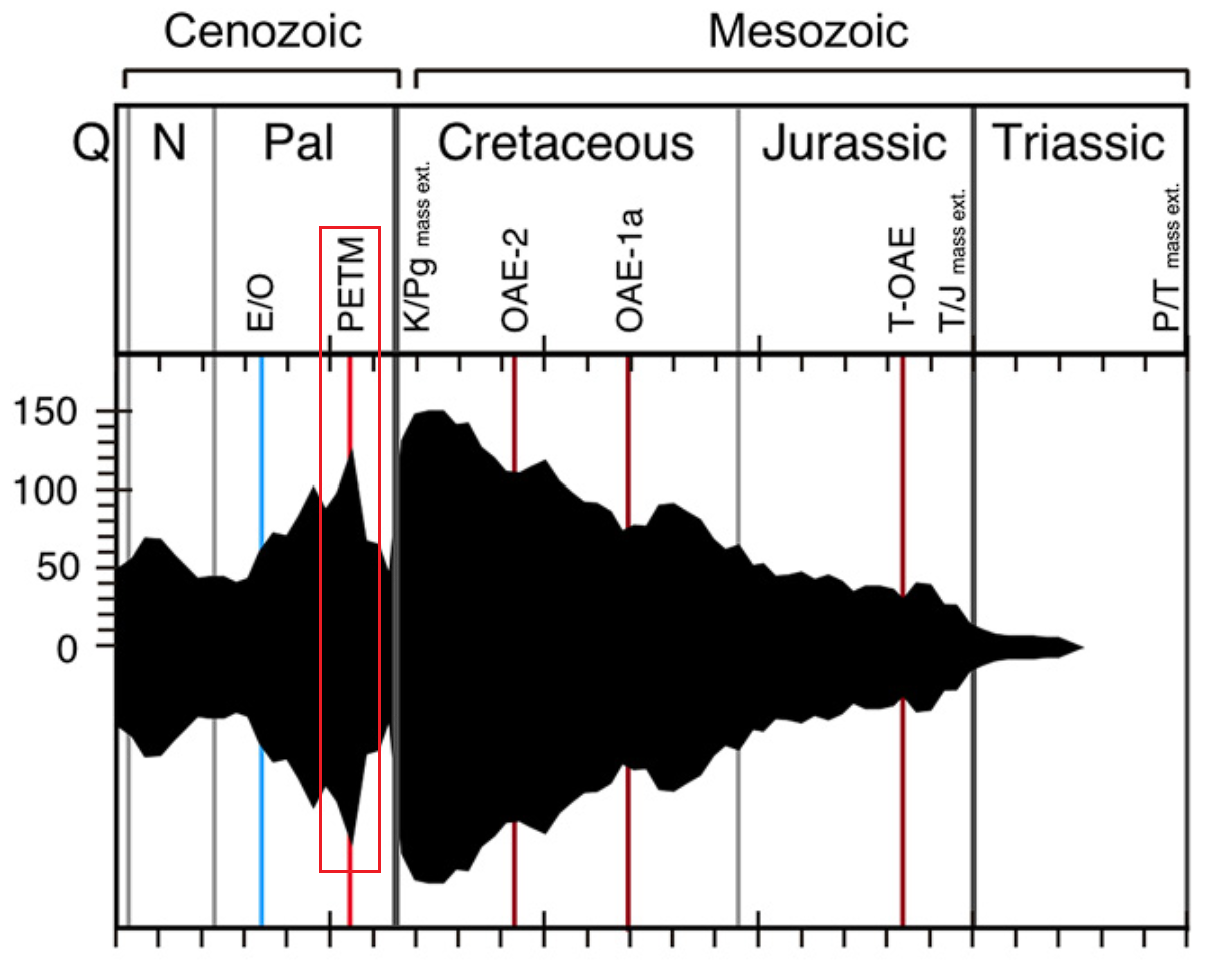
(上图:球石藻种类丰富程度随时间的变化;这张图的中生代部分其实上文中已经放过一遍了。请注意在 $\text{PETM}$ 期间的尖峰(红框框出部分))
这一“尖峰”的形成直到今天仍众说纷纭。许多证据表明,在 $\text{PETM}$ 期间,海洋同样经历了酸化和缺氧,而另一类用钙质构建外壳的生物——有孔虫(主要是底栖类群)同样遭遇了巨大的损失,直接灭绝了 $35\%-50\%$。这样一来,似乎球石藻的遭遇也算合理了。
之后的两千万年里,虽然有一些起伏,但总体来看球石藻的多样性一直在下跌。在距今约 $3400$ 万年前,发生了始新世-渐新世灭绝事件($\text{Eocene-Oligocene extinction event}$),而随着永冻冰盖再度从南北两极蔓延开来,球石藻再度遭到了重大的打击。球石藻目一半以上的属从此消失,合盤藻目的合盤藻科($\text{Zygodiscaceae}$)和等鞭金藻目的普氏石藻科($\text{Prinsiaceae}$)也至此绝灭。

(上图:第一排:合盤藻科的化石,从左到右分别是 $Lophodolithus$ 属、$Neochiastozygus$ 属、$Isthmolithus$ 属;第二排:普氏石藻科的化石,从左到右分别是:$Prinsius$ 属、$Toweius$ 属和 $Neobiscutum$ 属)
此后直至今日,球石藻在浮游藻类中的占比一直在 $15\%$ 左右,处于保三冲二的位置。平心而论,它们的成绩已经很好了,但作为一个曾经登顶过王者宝座的角色,如今它们的境遇总是有些差强人意。
所以,球石藻的故事就到此为止了吗?
尾章:诡谲的传奇
就在 $\text{PETM}$ 期间,一类前所未见的鳞片化石出现在了地层之中。它们形态修长、中空,开口处尖锐,仿佛是存放利剑的刀鞘一般,因此得名“剑球石”($\text{Gladioliths}$)。虽然这种化石始终极为稀少,反映出了其主人生存的窘境,但至少在几千万年间一直香火不绝,直至今日。这个神秘的类群被称为剑石藻($Gladiolithus$),而我们至今仍不知其所从来。分子生物学证据表明,它的演化位置可能处在钙鞭藻亚纲的最基部,但我们依旧难以想象它走过了怎样的演化道路,又为何能够熬过那么多次灭绝事件,最终在新生代涅槃重生:
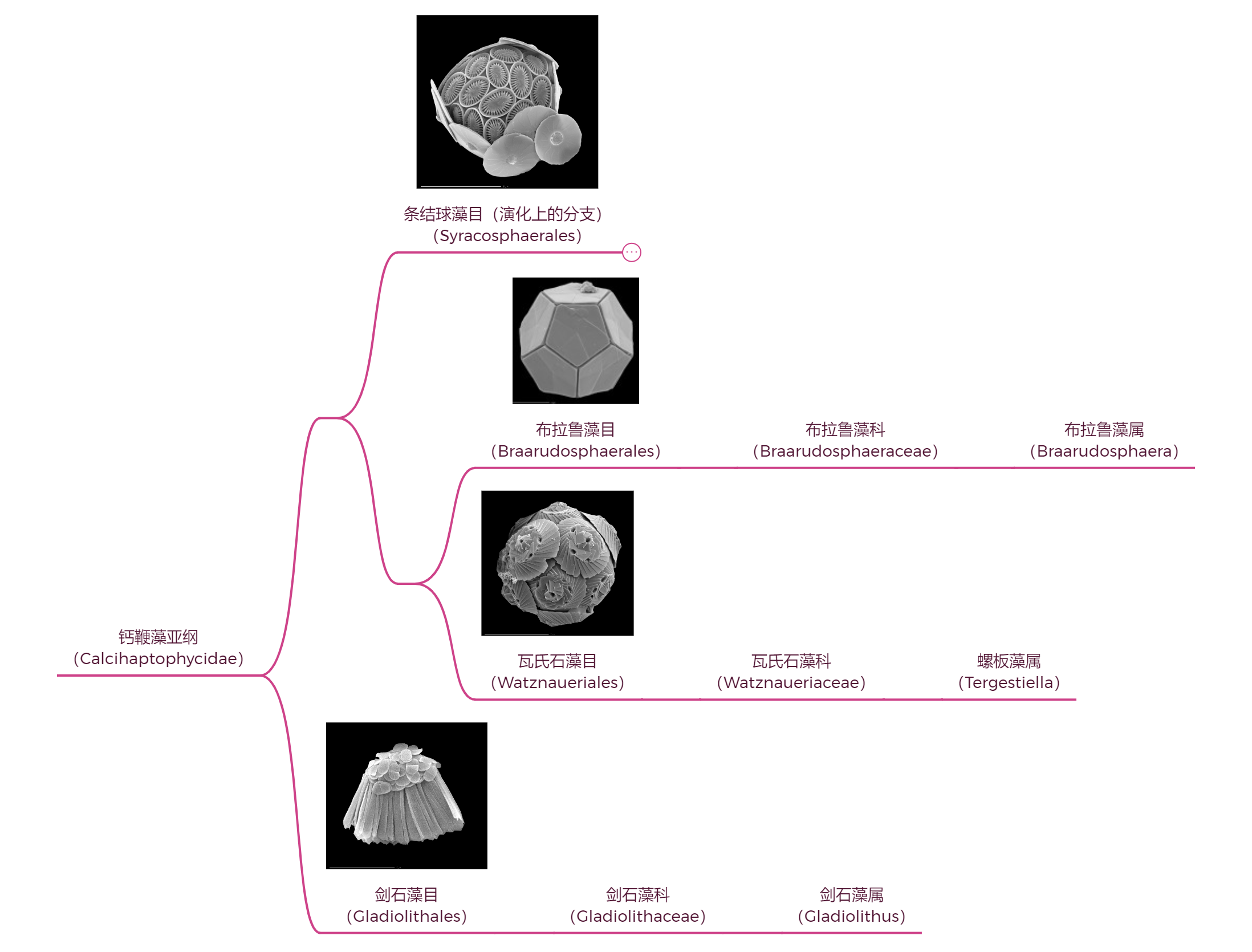
(上图:由分子生物学推定的钙鞭藻几大现存分支的演化树)
除了剑球石,剑石藻还有一种几乎没有任何修饰的椭圆形鳞片,怎么看都像是远古版本的遗物。这些鳞片构成了一套几乎密不透风的圆台形铠甲,乍看上去就仿佛是一块杯糕;在显微镜下,则能明显地看到许多竖直方向的脉络:
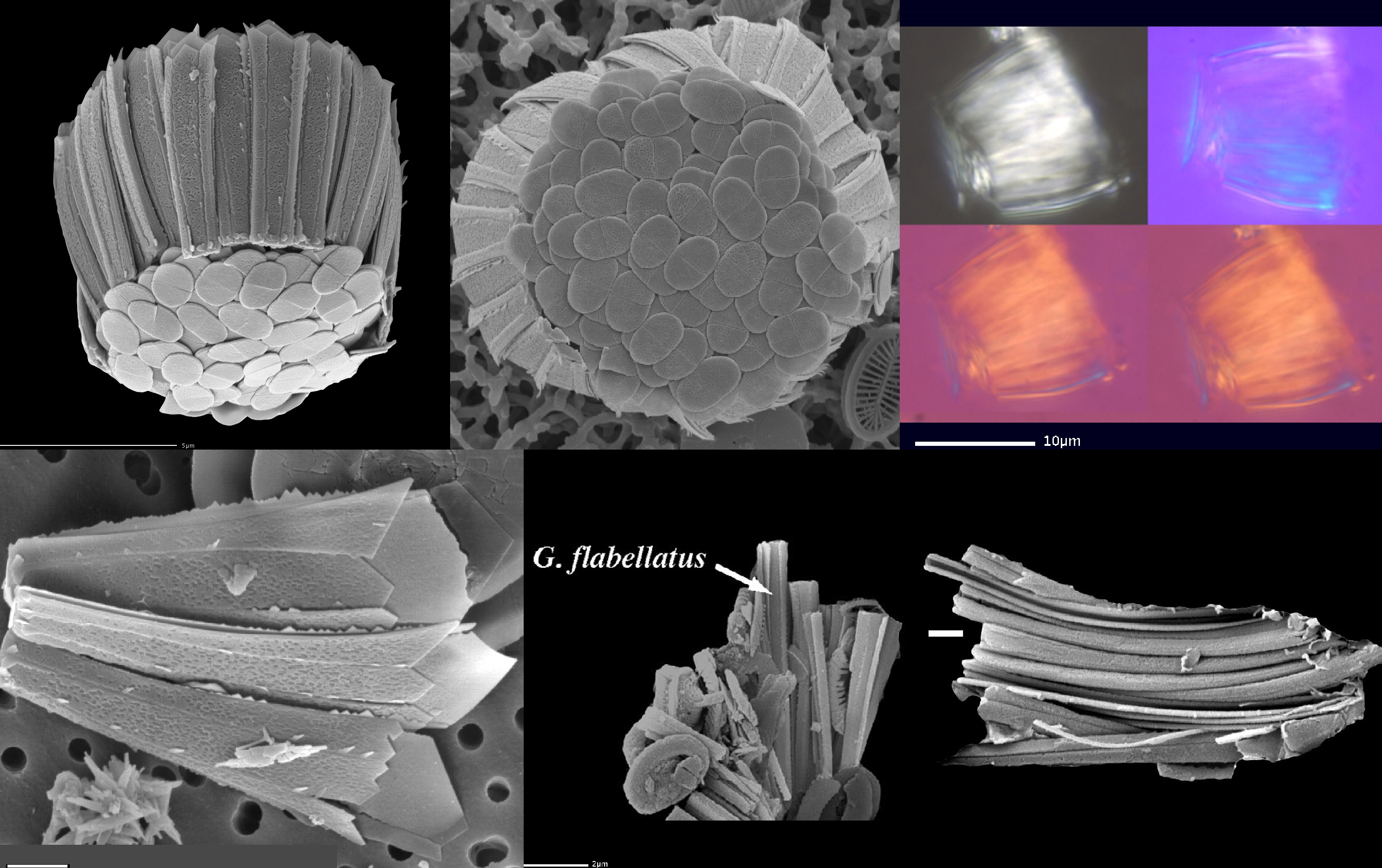
(上图:$G.~flabellatus$)
像剑石藻这样令人摸不着头脑的类群并不在少数。在始新世-渐新世灭绝事件后不久,地层中出现了一类马蹄形的鳞片化石,被称为“角球石”($\text{Ceratoliths}$)。古生物学家发现,即使不论这奇怪的形状,单看其晶体结构就与大部分钙鞭藻相去甚远,以至于都有人怀疑这货到底属不属于球石藻纲。后来,人们发现这类化石的主人——尖石藻($Ceratolithus$)并未灭绝,它依然存在于今日的海洋中;那种马蹄形的奇怪鳞片是它在单倍体阶段产生的,而其他几乎所有特征都表明它确实是钙鞭藻的一份子:
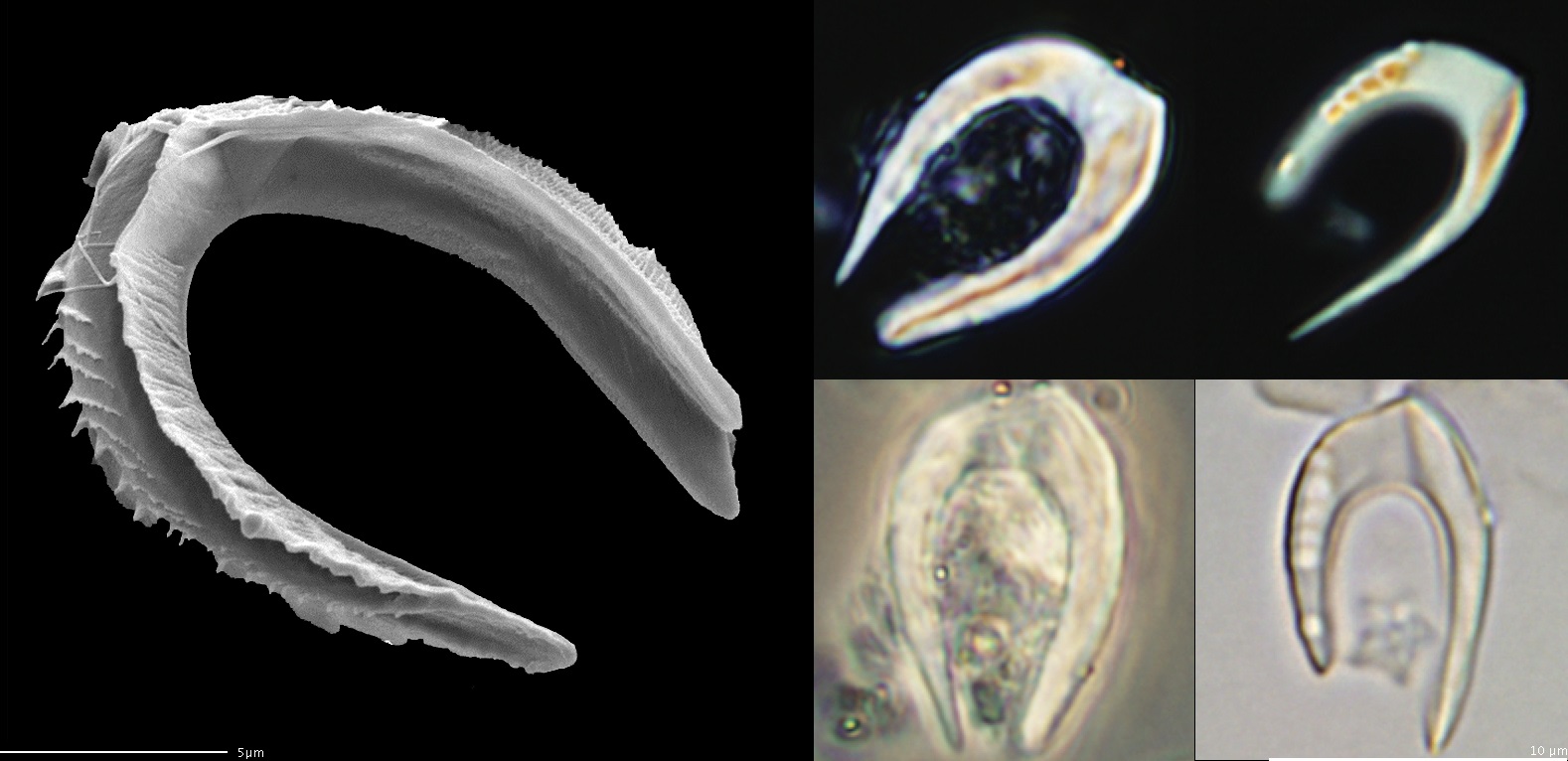
(上图:尖石藻产生的角球石。事实上,我们至今不知道尖石藻的单倍体阶段长成什么样。考虑到角球石的尺寸与形状,它似乎不是用于构建铠甲,那它到底有什么作用又是一个难以解释的问题)
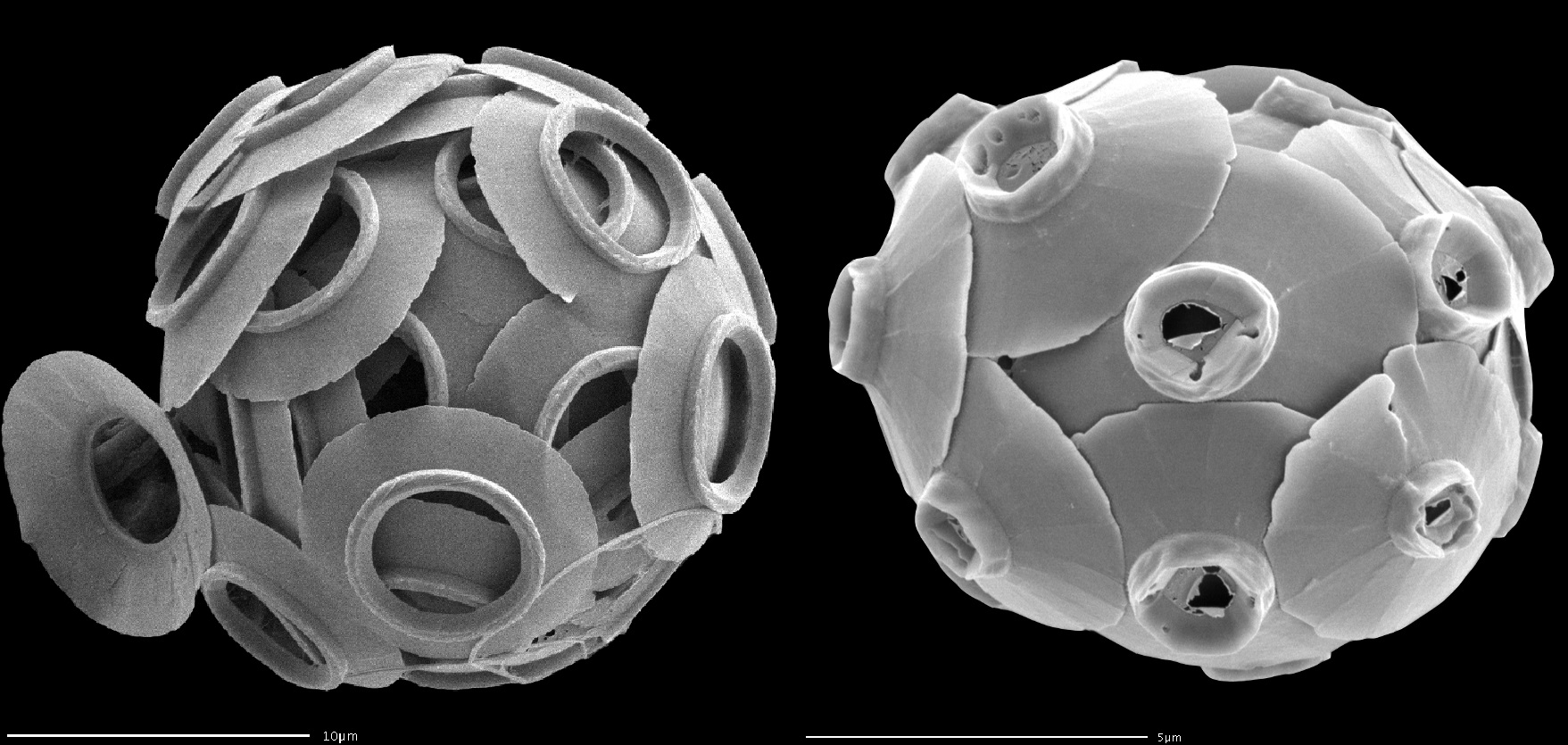
(上图:二倍体阶段的尖石藻 $C.~cristatus$(左)与 $C.~vidalii$(右))
如果说尖石藻好歹还有一些化石记录,那埃氏尖藻科($\text{Ericiolaceae}$)就几乎是凭空出现在了今天的海洋中,任凭古生物学家走遍四海、阅尽地层也找不到一丁点化石记录。科下共有两属,体型都奇小无比(仅 $5μm$ 左右),但埃氏尖藻属($Ericiolus$)却拥有两根长达十几微米的鞭毛,具有与一众钙鞭藻都格格不入的强悍运动能力;它的鳞片类似于三脚架,顶端异常尖锐,但只是松松垮垮地黏附在细胞表面。另一个帽球藻属($Pileolosphaera$)在外形上保持着普通钙鞭藻的模式,但其鳞片的几何对称性表明了它与埃氏尖藻的关系,也因此被归为同一科:
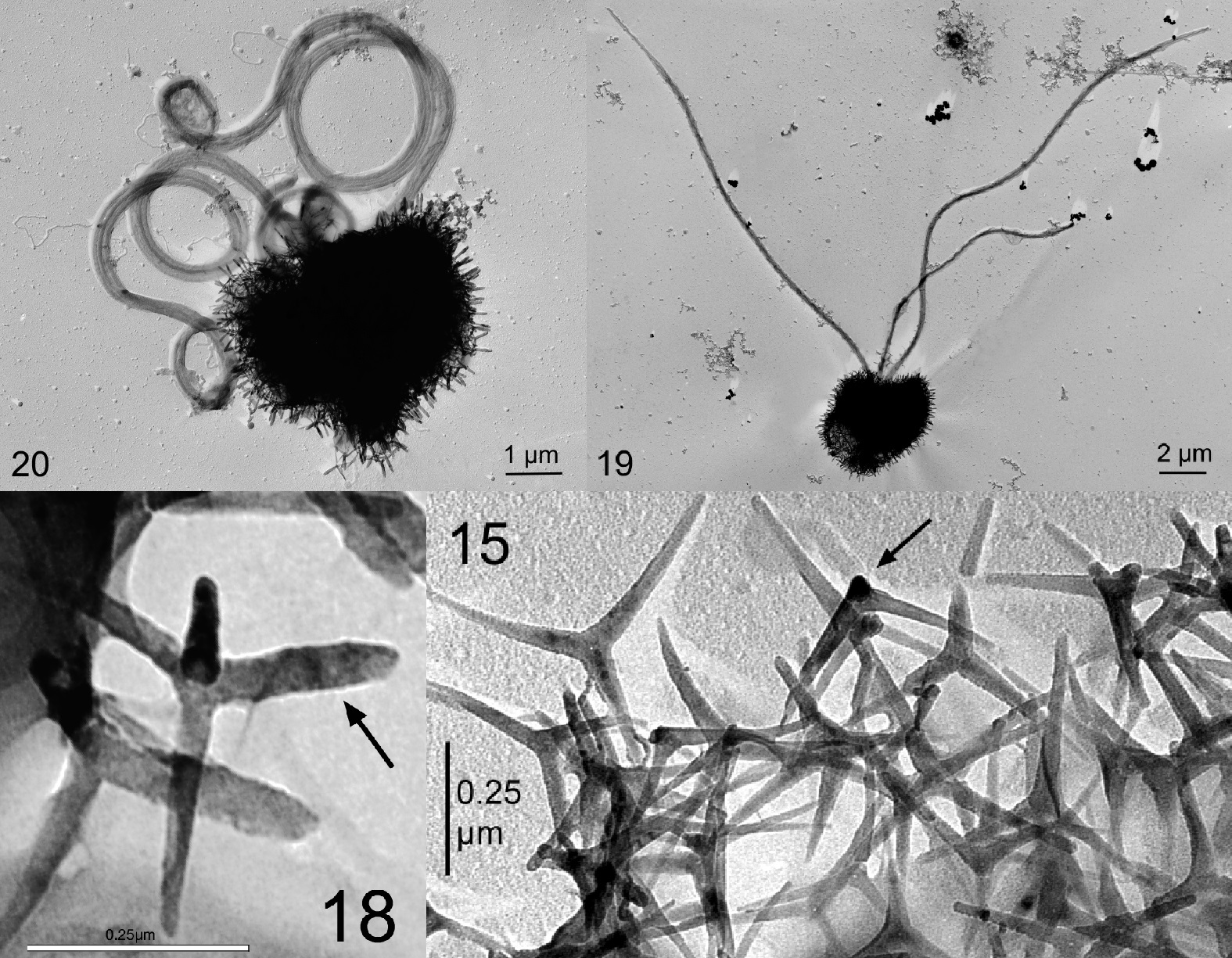
(上图:埃氏尖藻属的 $E.~pusillus$(第一行)及其鳞片(第二行))

(上图:帽球藻属的 $P.~longistirpes$。仔细看可以发现其鳞片也有三重对称性,与埃氏尖藻属很相似)
有些学者认为,埃氏尖藻科可能是盘星石目($\text{Discoasterales}$)的一支。这是一个在新生代中前期一度十分繁盛的钙鞭藻类群,鳞片呈放射状,拥有一个明显的中心;从这点看,二者确实很像。不幸的是,所有确认属于该目的物种都已灭绝,所以也很难验证该说法的正确性。
(上图:$D.~surculus$(属于盘星石属)的雪花状鳞片化石。它的灭绝($258.8$ 万年前)标志着新近纪的结束,第四纪的开始)
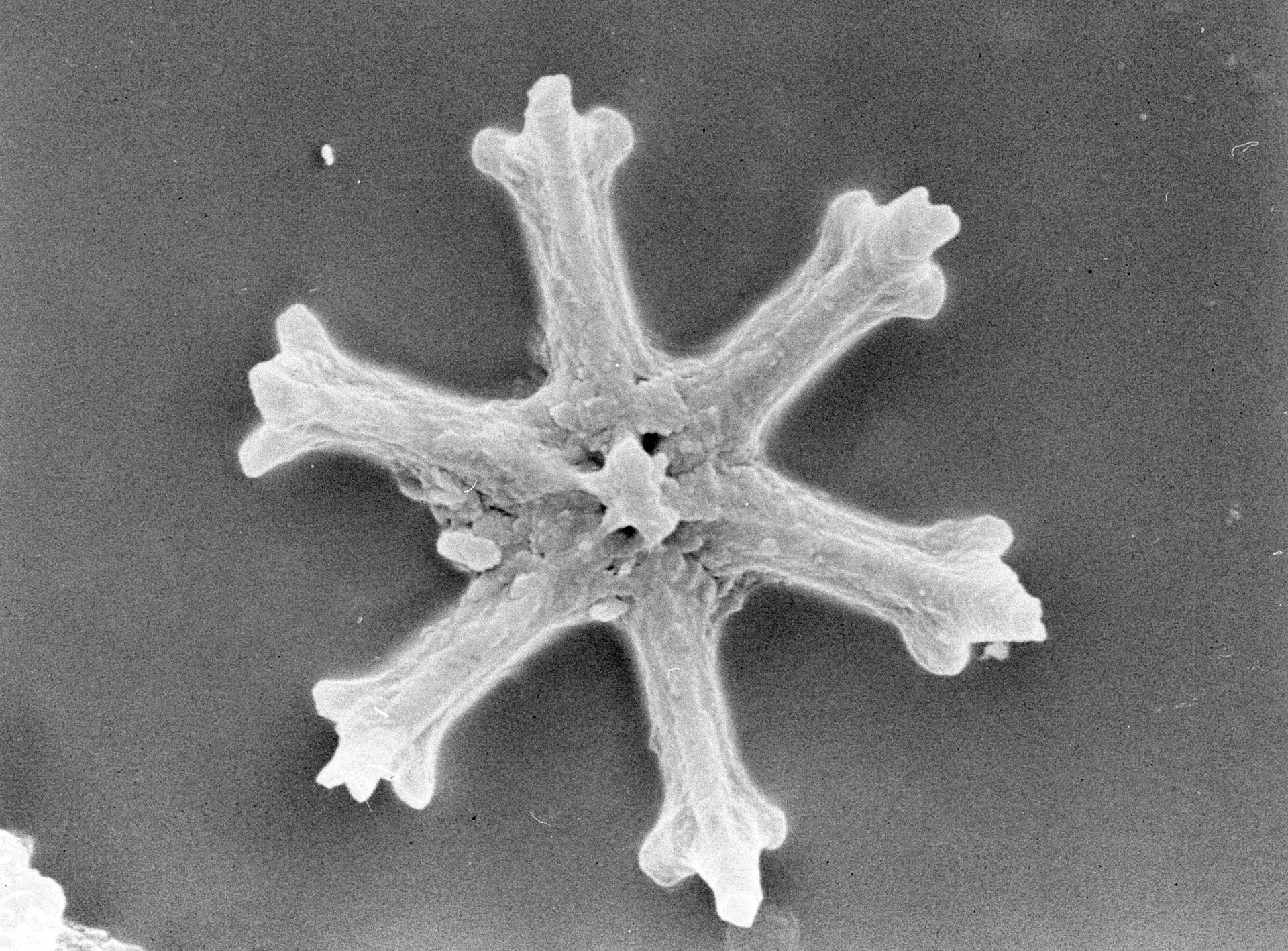
如果你觉得埃氏尖藻已经很奇怪了,那请别急,更加抽象的还在后面。在条结球藻目下,还有一个长相离了大谱的类群,那就是迈氏球藻亚科($\text{Michaelsaroidae}$)。其掌门人迈氏球藻($Michaelsarsia$),乍看起来仿佛是一只有着十几条大长腿的蜘蛛,每条腿还分为 $2-3$ 节,若不是有旁边的比例尺,恐怕还真有人要把它当成什么节肢动物。若细细观之,你会发现那些“腿”(专业点说叫附球石,$\text{appendage coccolith}$)不过是由一种特化的鳞片组装起来的罢了,只能起到支撑或者防御的作用,而不能真正地运动起来:
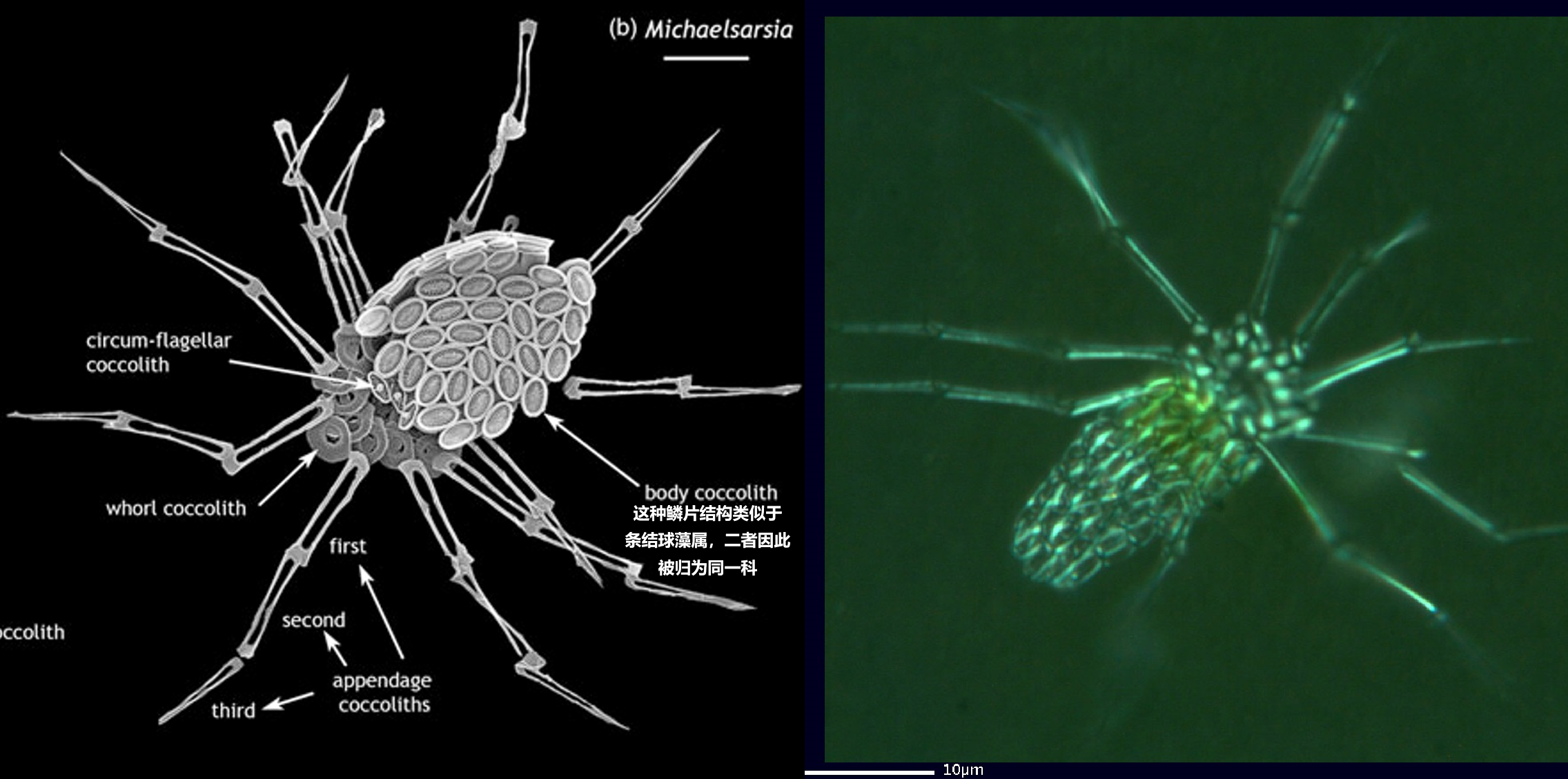
(上图:迈氏球藻属的外壳结构(左)与 $M.~adriaticus$(右))
同亚科的近亲蛇星藻($Ophiaster$)更是奇形种。它的本体之外有好几条类似于触手的附球石,长度可达几十微米,而细细观之,你会发现它们也是一节一节的,由某种特化的鳞片拼装而成。“触手”从铠甲背部一点出发,有时几乎是一条直线,有时却又极尽弯曲之能事,如蛇一样盘绕。这么看,“蛇星藻”这个名字真是形象至极:
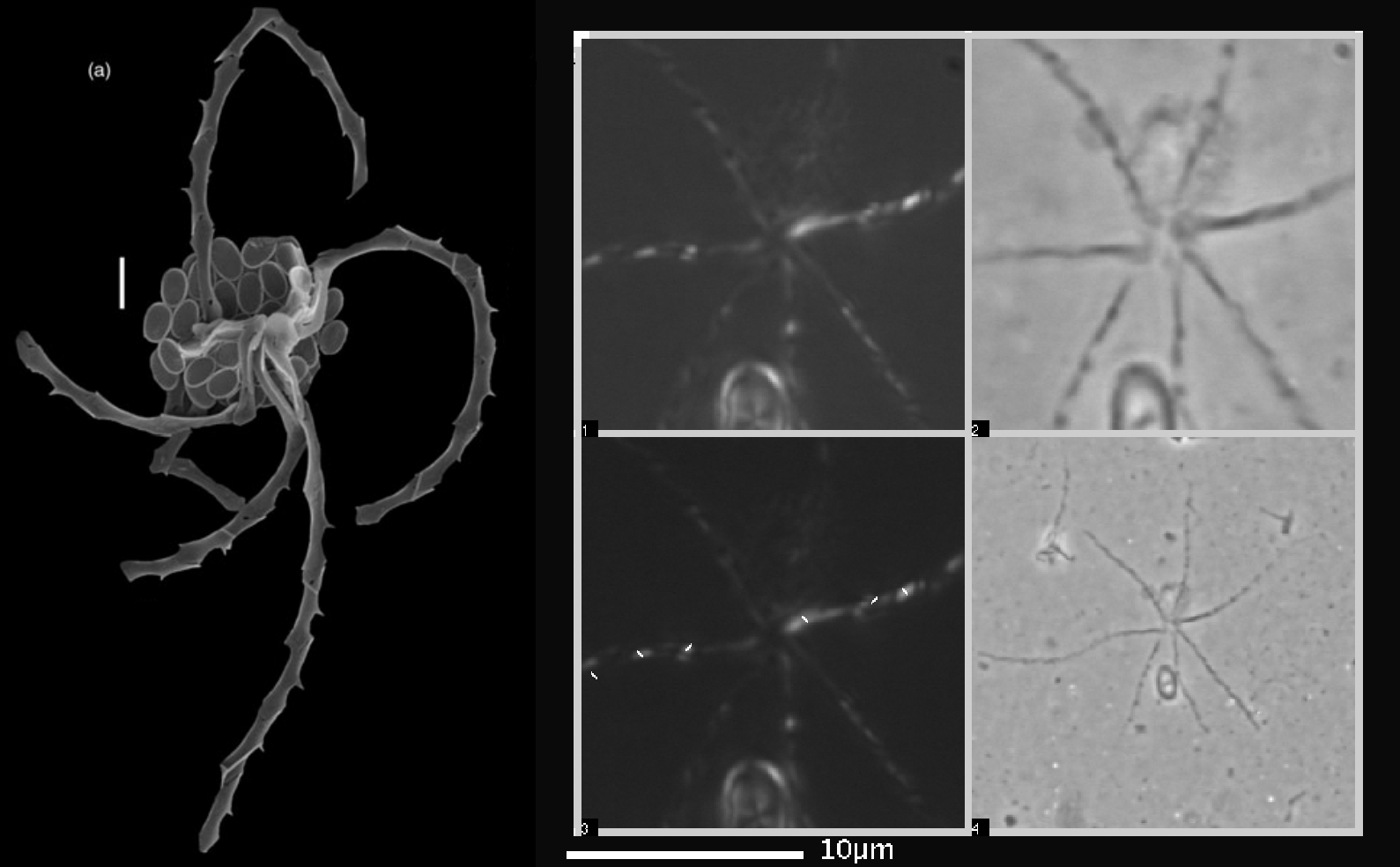
(上图:$O.~hydroideus$ 标本(左)与某种蛇星藻的照片(右,具体物种不明))
当然,迈氏球藻之流也绝非抽象的极限,因为在我看来,现存形态最诡异的钙鞭藻非伞球藻($Umbellosphaera$)莫属。它的鳞片确实像一把撑开的伞,但因为中央凹陷,所以看起来更像是喇叭或者漏斗;这些“伞”以一种超越想象的方式排布,最终构建起它那套简直不可名状的铠甲,让我深刻地体会到了人类语言的无力:

(上图:$U.~irregularis$(左)与 $U.~tenuis$(右))

(上图:$U.~irregularis$)
从化石记录上看,伞球藻是在第四纪($\text{Quaternary}$)早期(约 $250$ 万年前)突然出现的,而其离谱的形态又让人难以将它和其他类群联系起来。不过,最近的一项分子生物学研究表明,伞球藻可能是棒球藻科的姐妹,但支持率并不高,难以服众。只能说,在球石藻的演化上,还有众多的谜团等待我们去解决。
补充与讨论
我们在上文中已经讲述了许多关于定鞭藻的演化故事,但需要明确的是,由于任何生物都无法主动选择自己的演化方向,所以这些故事并不严谨。演化的本质是物竞天择,适者生存,但若真的像这样描述每一个演化细节未免过于啰嗦,所以上文(以及下文)中还是采用了类似于它们在主观选择的笔调,实为无奈之举。
整体来看,定鞭藻还是众多藻类中比较成功的一支,即便如今正处于演化史上的低谷,它们的身影依旧遍布海洋与淡水的整个阳光带,是浮游生物的重要组成部分。球石藻作为门内最繁盛的一支,对于海洋生态系统的贡献尤为巨大。超过 $50\%$ 的海洋中上层 $\ce{CaCO3}$ 沉积来自于它们,以至于当球石藻大量繁殖之时,整个海洋的反照率都会因此改变(前文已有提到);所谓的“大方解石带”($\text{Great Calcite Belt}$)也是由它们形成:
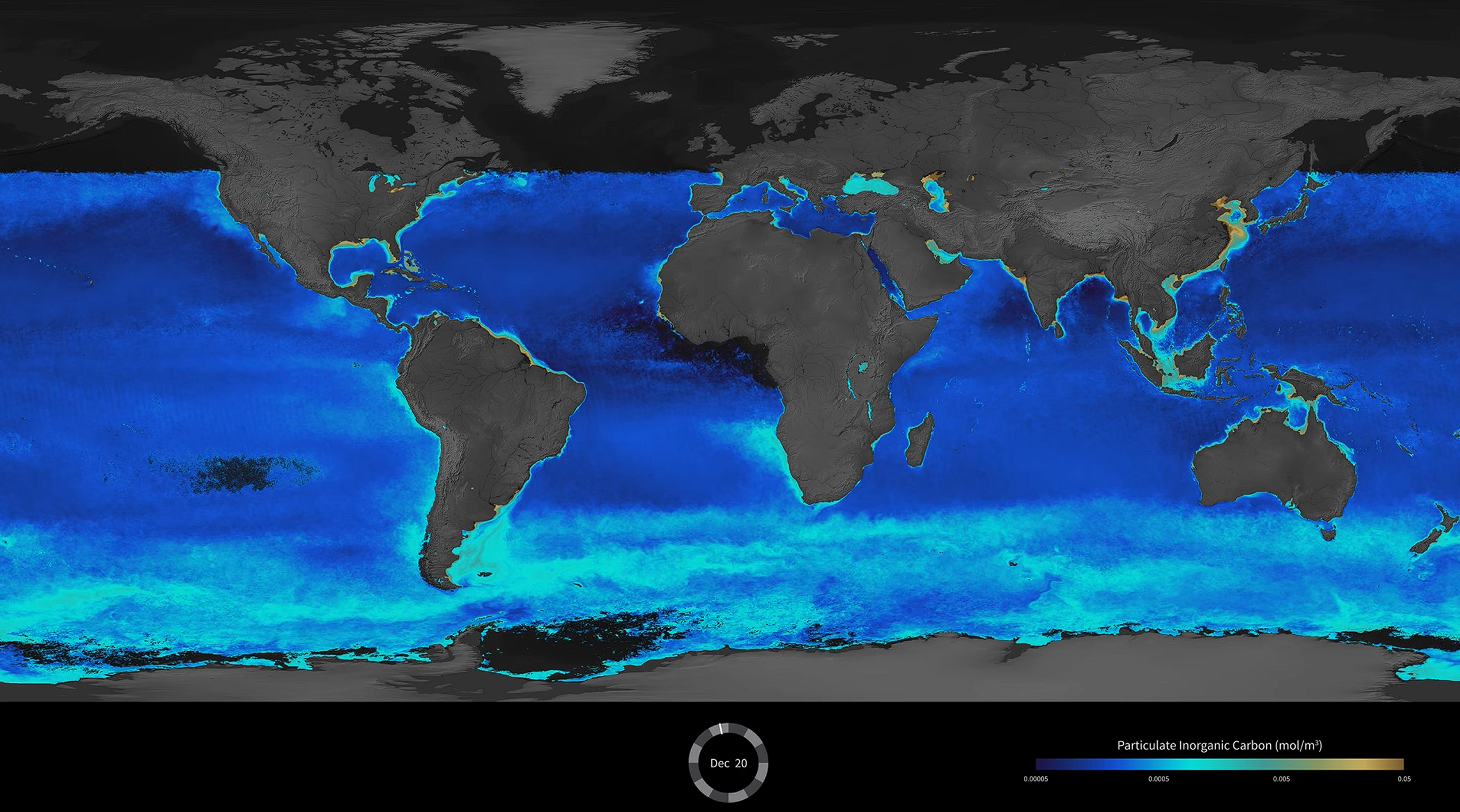
(上图:冬季全球海域无机碳颗粒密度。可以看到横贯南大洋的一条亮带,即是所谓“大方解石带”)
由于这些 $\ce{CaCO3}$ 最终会沉入海底,形成白垩,所以球石藻用于制造铠甲的那部分碳会因此脱离生物圈,并在短时间内难以重返。它们就像是无数台微小的抽气机,在亿万年的时光里一直忠实地履行着自己的使命,将难以想象的庞大数量的二氧化碳拖入海底,最终被地层封印。若此时再回看那壮观的多佛白崖,你会觉得那并不是球石藻的坟墓,而是它们永恒的丰碑。
当然,球石藻在地层中留下的还不止于此。它们的生命活动中会产生一类叫做长链烯酮($\text{Long-chain Alkenones}$)的独特分子,而大量研究表明,当外界环境改变时,它们产生的长链烯酮结构也会随之变化。因此,通过分析沉积物中的这类物质,我们便可以反推出那一时代的环境。于是,在三叠纪末牙形石(牙形动物纲,$\text{Conodonta}$)灭绝之后,球石藻留下的长链烯酮以及其他新一代生物在地层中留下的痕迹就这样无缝衔接了它的使命,继续书写着地球环境的变迁:
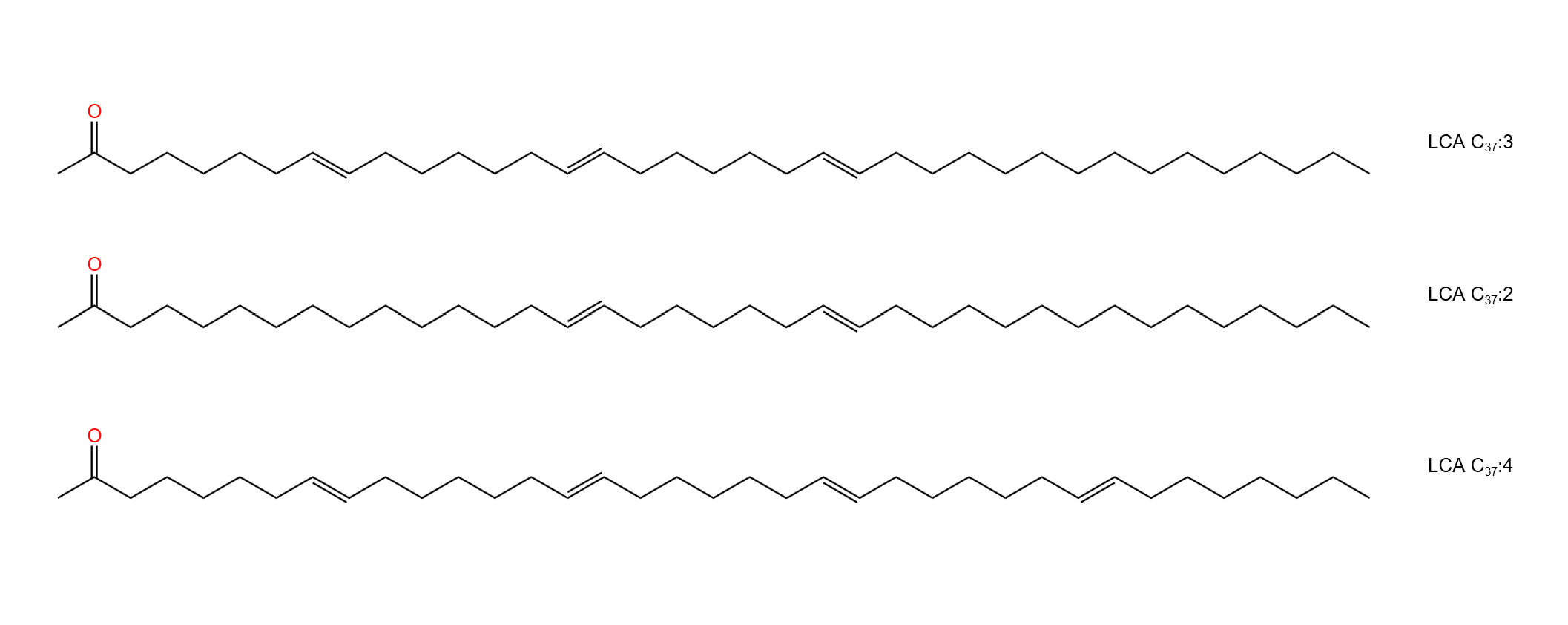
(上图:三种典型的长链烯酮。右侧的编号中,$\ce{C37}$ 表示有 $37$ 个碳原子,冒号后面的表示双键个数)

(上图:牙形石,一种磷灰石质的微小化石。大部分牙形石被确认属于一类脊椎动物,由于其数量极为丰富且具有一些特性,所以被广泛用于古地质学研究。牙形石在三叠纪末大灭绝后消失)
如果跳出地层,你还会发现这些小东西的另一项丰功伟绩。不知道各位读者在去海边时有没有闻到过“大海的气味”,有些人觉得这种味道对身体有益,但也有些人觉得这是一种难以言表的恶心臭味。最早,人们认为这一气味来自于臭氧($\ce{O3}$),但到了二十世纪中期,科学家最终确认这种气味来源于二甲基硫醚($\ce{(CH3)2S}$,$\text{DMS}$)。说到这里,有些人会想起不久前,一群天文学家宣称他们在一颗系外行星($\text{K2-18b}$)上发现了生命可能存在的迹象,其实就是他们在这颗行星的大气中检测出了 $\text{DMS}$ 的存在。那么,这种物质又为何能成为生命的标志呢?因为至少在地球上,$\text{DMS}$ 只能由生命合成,而任何非生命的过程,无论是雷电、火山还是其他什么都无法产生 $\text{DMS}$。那么,又到底是那类生物合成了这种物质呢?
严格来说,有很多类生物共同参与了这一过程,但其中的主力正是定鞭藻。具体来说,定鞭藻会在胞内合成一种叫 $\text{DMSP}$($\ce{(CH3)2S+CH2CH2COO−}$)的物质,用于调节渗透压、维持细胞形态,而当这种物质因为各种原因(比如细胞死亡)被释放到外界环境中后,就会被细菌等微生物分解,产生挥发性的 $\text{DMS}$。最后,一部分 $\text{DMS}$ 进入大气,让你闻到了“大海的气味”:
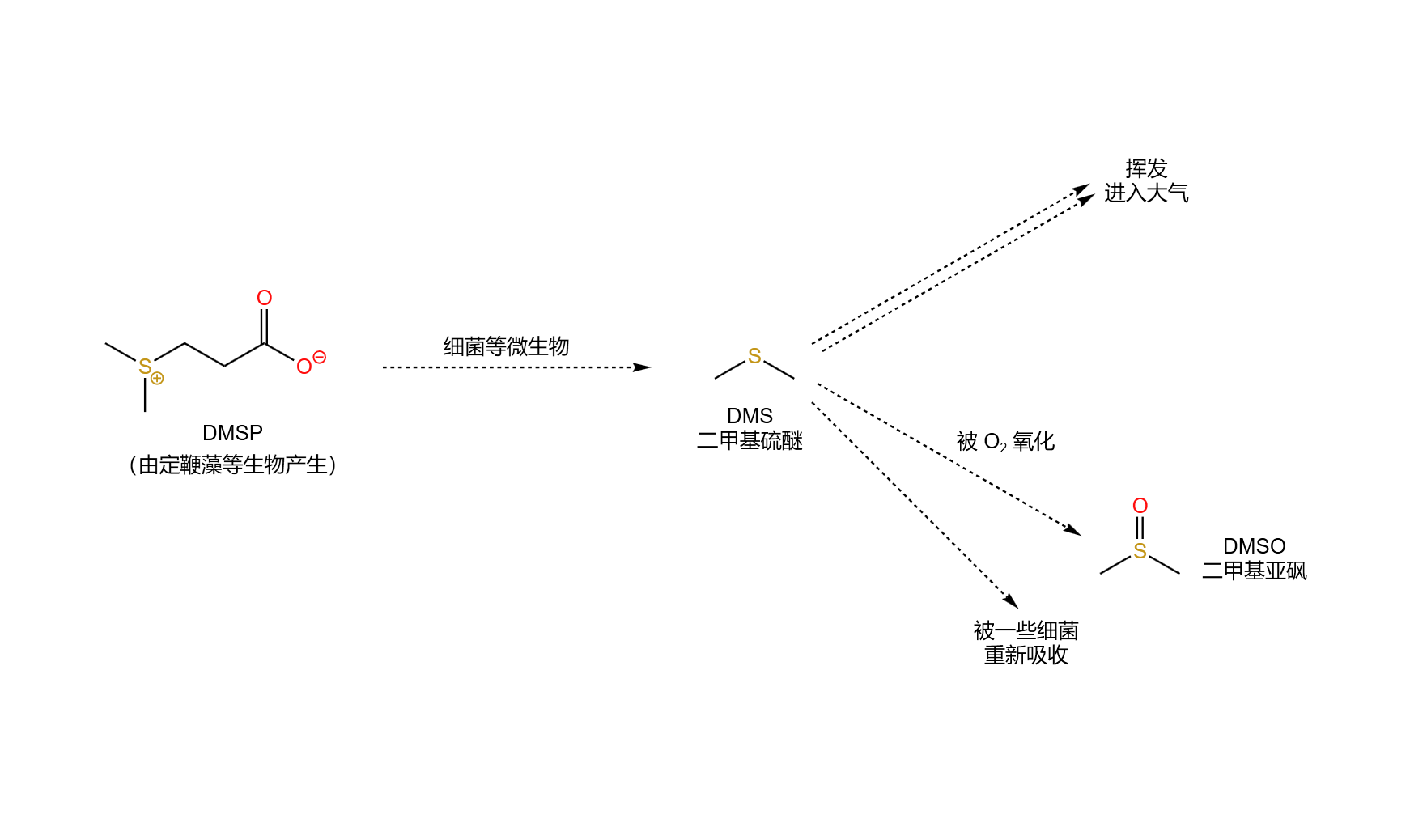
(上图:有关 $\text{DMS}$ 的生物过程。请注意,这幅图以及上文的描述是经过简化的,实际情况要复杂得多,有兴趣的读者可以自行阅读相关文献)
所以,这一过程只是为了让我们闻个味?当然不是。大量研究表明,$\text{DMS}$ 对于地球气候有重要影响,这就不得不提到著名的CLAW假说。该假说认为,$\text{DMS}$ 在大气中会被氧化,由此产生的一系列物质将形成气溶胶,并充当凝结核促使云和降雨的生成。当全球变暖、定鞭藻大量繁殖之时,更多的 $\text{DMS}$ 随之产生,从而导致云量增加、到达地表的阳光减少,进而使得全球变冷,由此形成的负反馈循环可能在上亿年的时光中保证了地球气候的稳定。
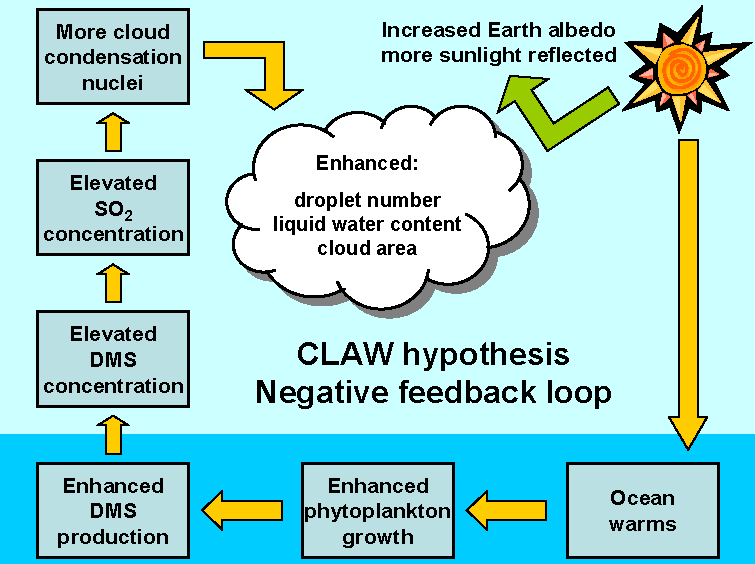
(上图:$\text{CLAW}$ 假说的示意图。值得注意的是,这套负反馈体系是可以双向运行的,即若全球变冷,一套流程下来会导致到达地表的阳光增加,进而使得全球变暖。还有一些研究者认为 $\text{CLAW}$ 假说的模型过于简单,$\text{DMS}$ 实际对气候的影响可能更大,涉及的过程也可能更加复杂)
写到这里,我突然感觉此前讲述的故事都有些自作多情了。在演化的修罗场上,胜负成败本就是日常之事,或许它们本就不在乎这一城一池的得失;但是,在气候的调节、生态系统的构建中,这个不知名的渺小族裔却倾尽全力,在大多数人都不知道的角落默默地守护着天下苍生。无论是否身处王者之位,它们都始终坚守着那份使命,而只要使命仍在,定鞭藻就不会走向末路。
后记
终于把这篇文章写完了!我敢说,这应该是中文互联网上最全面、最细致的梳理定鞭藻演化脉络的文章。事实上,由于分子生物学证据的匮乏和混乱,如今学术界对于定鞭藻的演化也颇有争议,所以上文中的故事很多都只是一家之言,但对于科普文章来说,或许也只能如此。当然,本人并不是此方面的专业人士,只是作为爱好者的心血来潮罢了,所以若有什么错误或遗漏之处,还敬请各位指出,在此感谢!
最后还是老样子,贴上我整理的定鞭藻门系统发育树:
(好吧,图片依旧不清楚)以及它的 $\text{pdf}$ 版链接:Link
参考资料
网站
$[1]$:Jeremy R. Young et al., Nannotax 3, an authoritative guide to the biodiversity and taxonomy of coccolithophores. Searched on July 17, 2025. Link(这是一个非常好的网站,由此领域的泰斗主持编写,旨在为球石藻的分类学提供权威指南,同时收录了上万张球石藻的照片,包括现生的和灭绝的物种;网站还整理了时间轴,标明了各种化石在地层中何时出现、何时消失,这对理解球石藻的演化大有帮助。大力推荐所有球石藻爱好者浏览该网站)
$[2]$:The International Nannoplankton Association, an association of scientists worldwide who have an interest in living or fossil nannoplankton. Searched on July 17, 2025. Link(国际微浮游生物联合研究会官网)
$[3]$:WoRMS - World Register of Marine Species, an authoritative classification and catalogue of marine names. Searched on July 17, 2025. Link(世界海洋物种目录官网,是一个致力于提供全面的、具有公信力的海洋物种目录的权威生物学数据库)
$[4]$:AlgaeBase - Listing the World’s Algae, a global algal database of taxonomic, nomenclatural and distributional information. Searched on July 17, 2025. Link(AlgaeBase 是一个收录世界各地陆生、淡水与海洋藻类与海草物种及其照片的权威数据库)
论文
$[5]$:Parfrey L W, Lahr D J G, Knoll A H, et al. Estimating the timing of early eukaryotic diversification with multigene molecular clocks[J]. Proceedings of the National Academy of Sciences, 2011, 108(33): 13624-13629.
$[6]$:Penot M, Dacks J B, Read B, et al. Genomic and meta-genomic insights into the functions, diversity and global distribution of haptophyte algae[J]. Applied Phycology, 2022, 3(1): 340-359.
$[7]$:Yang P, Guo K, Yang Y, et al. Phylogeny and genetic variations of the three genome compartments in haptophytes shed light on the rapid evolution of coccolithophores[J]. Gene, 2023, 887: 147716.
$[8]$:Kawachi M, Nakayama T, Kayama M, et al. Rappemonads are haptophyte phytoplankton[J]. Current Biology, 2021, 31(11): 2395-2403. e4.
$[9]$:Véron B, Rougier E, Taylor A, et al. New species of Pavlovophyceae (Haptophyta) and revision of the genera Exanthemachrysis, Rebecca and Pavlova[J]. European journal of taxonomy, 2023, 861: 21-47.
$[10]$:Andrews H T, Wujek D E. Ecology, Reproduction, and Classification of the Haptophyte Alga Apistonema From Kansas[J]. The Southwestern Naturalist, 2017, 62(4): 292-295.
$[11]$:Edvardsen B, Eikrem W, Throndsen J, et al. Ribosomal DNA phylogenies and a morphological revision provide the basis for a revised taxonomy of the Prymnesiales (Haptophyta)[J]. European journal of phycology, 2011, 46(3): 202-228.
$[12]$:Wang X, Fon M, Andersen A J C, et al. Insights into the nature of ichthyotoxins from the Chrysochromulina leadbeateri blooms in Northern Norwegian fjords[J]. Harmful Algae, 2024, 137: 102681.
$[13]$:Gjøsæter J, Lekve K, Stenseth N C, et al. A long-term perspective on the Chrysochromulina bloom on the Norwegian Skagerrak coast 1988: a catastrophe or an innocent incident?[J]. Marine Ecology Progress Series, 2000, 207: 201-218.
$[14]$:Fallon T R, Shende V V, Wierzbicki I H, et al. Giant polyketide synthase enzymes in the biosynthesis of giant marine polyether toxins[J]. Science, 2024, 385(6709): 671-678.
$[15]$:Nakayama T, Yoshida M, Noël M H, et al. Ultrastructure and phylogenetic position of Chrysoculter rhomboideus gen. et sp. nov.(Prymnesiophyceae), a new flagellate haptophyte from Japanese coastal waters[J]. Phycologia, 2005, 44(4): 369-383.
$[16]$:Monteiro F M, Bach L T, Brownlee C, et al. Why marine phytoplankton calcify[J]. Science Advances, 2016, 2(7): e1501822.
$[17]$:Strom S L, Bright K J, Fredrickson K A, et al. Phytoplankton defenses: Do Emiliania huxleyi coccoliths protect against microzooplankton predators?[J]. Limnology and Oceanography, 2018, 63(2): 617-627.
$[18]$:Lewis W H, Paris G, Beedessee G, et al. Plastid translocon recycling in dinoflagellates demonstrates the portability of complex plastids between hosts[J]. Current Biology, 2024, 34(23): 5494-5506. e3.
$[19]$:Coale T H, Loconte V, Turk-Kubo K A, et al. Nitrogen-fixing organelle in a marine alga[J]. Science, 2024, 384(6692): 217-222.
$[20]$:Scotese C R, Song H, Mills B J W, et al. Phanerozoic paleotemperatures: The earth’s changing climate during the last 540 million years[J]. Earth-Science Reviews, 2021, 215: 103503.
$[21]$:Taylor A R, Chrachri A, Wheeler G, et al. A voltage-gated H+ channel underlying pH homeostasis in calcifying coccolithophores[J]. PLoS biology, 2011, 9(6): e1001085.
$[22]$:Meyer E M, Taylor A R. A comparison of calcification mechanisms in haploid and diploid cells of the coccolithophore Calcidiscus leptoporus (Murray & Blackman 1898)[J]. Journal of Phycology, 2025, 61(3): 466-484.
$[23]$:Sáez A G, Probert I, Young J R, et al. A review of the phylogeny of the Haptophyta[J]. Coccolithophores: From molecular processes to global impact, 2004: 251-269.
$[24]$:Bown P R, Young J R, Lees J A. On the Cretaceous origin of the Order Syracosphaerales and the genus Syracosphaera[J]. Journal of Micropalaeontology, 2017, 36(2): 153-165.
$[25]$:Gran‐Stadniczeñko S, Šupraha L, Egge E D, et al. Haptophyte diversity and vertical distribution explored by 18S and 28S ribosomal RNA gene metabarcoding and scanning electron microscopy[J]. Journal of Eukaryotic Microbiology, 2017, 64(4): 514-532.
$[26]$:Young J R, Andruleit H, Probert I. COCCOLITH FUNCTION AND MORPHOGENESIS: INSIGHTS FROM APPENDAGE‐BEARING COCCOLITHOPHORES OF THE FAMILY SYRACOSPHAERACEAE (HAPTOPHYTA) 1[J]. Journal of Phycology, 2009, 45(1): 213-226.
$[27]$:Hagino K, Young J R, Morono Y. Molecular phylogenetic affinity of Hayaster cf. perplexus based on 18S rDNA sequences[J]. Journal of Nannoplankton Research, 2023, 41(2): 98-104.
$[28]$:I.T. Marlowe, J.C. Green, A.C. Neal, S.C. Brassell, G. Eglinton & P.A. Course (1984) Long chain (n-C37–C39) alkenones in the Prymnesiophyceae. Distribution of alkenones and other lipids and their taxonomic significance, British Phycological Journal, 19:3, 203-216.
$[29]$:Madhusudhan N, Constantinou S, Holmberg M, et al. New Constraints on DMS and DMDS in the Atmosphere of K2-18 b from JWST MIRI[J]. The Astrophysical Journal Letters, 2025, 983(2): L40.
$[30]$:Park J Y, Song J, Kim K Y, et al. Optimised primer set for accelerated amplification of RuBisCO large subunit (rbc L) gene and identification of haptophytes[J]. Journal of Applied Phycology, 2025: 1-10.
$[31]$:孙军, 靳少非. 中国近海今生颗石藻物种多样性初步研究[J]. 生物多样性, 2011, 19(6): 787-797.